2781
Views & Citations1781
Likes & Shares
BACKGROUND
Measles is caused by
the measles virus, a single-stranded, negative-sense, enveloped RNA virus.
Humans are the natural hosts of the virus; no other animal reservoirs are known
to exist. Monkeys in the wild apparently are free of measles, contracting infection
only after contact with humans [1], causes a highly contagious viral infection,
with typical clinical symptoms including maculopapular rash, fever, cough,
coryza and conjunctivitis. Measles continues to be a major cause of childhood
morbidity and mortality worldwide. Measles, which is an acute systemic
infection of, in most cases, young children ,is responsible for 10% of deaths
from all causes among children less than 5 years old [2].
In the early 1960s,
the advent of a live attenuated measles virus (MV) vaccine reduced the
incidence of measles in many parts of the world, including developed and
developing countries. In some developing countries, fatality rates for measles
can still be as high as 15%, causing about 770, 000 deaths among infants and children
and this is probably due to lack of vaccination of many individuals in the
population [3].
Although the introduction of live attenuated
MV vaccines had largely reduce the endemic circulation of wild-type MV in the
industrialized world, vaccination has been less successful in large areas of
Africa and Asia [4]. Elimination of MV requires increasing the vaccination
coverage levels, In this context, it is very important that reliable and
sensitive laboratory methods are used to accurately determine the antibody
level and protection achieved after vaccination and the level of antibodies
that persists in those who were previously vaccinated [5].
It is likely that many factors contribute to
the presence of susceptible individuals among mass vaccinated populations.
These include failure to seroconvert and decline of immunity with time after
vaccination [6]. Other important factors that might affect the immune response,
the age at the time of vaccination, the number of doses, and the strain
included in the vaccine [7]. Measles vaccination is not effective in very young
infants, mainly due to interference with MV-specific maternal antibodies and
immaturity of the immune system. In most industrialized countries with low
measles incidence the first measles vaccination is, therefore, carried out
between the ages of 12 and 15 months [8]. The protective immunity induced by
vaccination may not be life long without being boosted by an exposure, mostly
sub clinically, to a naturally circulating virus [9].
One year after the vaccination, 16% of the
children who received the vaccination were no longer protected [10]. More than
70% of children who received the first vaccination before 10 months of age were
shown to have NT antibodies at 15 months or older and that 98% of children who
received the second dose at this timing had measurable NT antibodies 8 months
after revaccination [11].
MATERIALS AND
METHODS
Study area
This study was conducted in three localities
in Khartoum state during the period from February to August 2014.
Study population
Children between the ages of 7 months and 7
years were included in this study. Details of the vaccination history were
obtained from there parental, concerning, date of vaccination and the type of
vaccine given.
Inclusion and
exclusion criteria
The children were selected based on the
following criteria: age; number of vaccine doses and no past history of severe
illnesses.
Sample size and
sampling method
One hundred blood samples were collected
based on the compliance of criteria, seventy three of them were subjected to
Neutralization test and 20/100 samples subjected to hemagglutinition inhibition
assay
Study design
This is an investigatory study and it
embraced children at different age group (7-9 months) – (11-15mounths) – (19-22
months) and (6-7years). Three localities were chosen for this study (Bahri,
Khartoum and Omdurman) and this was done to compare the immune status of
children in different parts after different number of vaccinations.
Ethical
considerations
The study was approved by Ministry of Health
and the hospital from which samples were collected and verbal consent was
obtained from subjects enrolled in the study.
Blood samples
collection, transport, processing and storage
3 ml of blood were collected in plain vacuum
tubes through venipuncture. The blood was kept for 2 h at 25°C then centrifuged
at 3000 rpm for 10 min. Serum was carefully removed with a Pasteur pipette and
transferred to sterile labeled cryovials, then stored at -20°C until used.
Source of vaccine
virus
The vaccine strain was obtained from the
Public Health Centralized Immunization Clinic. It was manufactured by the Serum
Institute of India Ltd. (Pune, India).
Virus propagation
The measles virus stock was propagated in a
vero cell line in two T.C 75 cm2 flasks, which were inoculated with
0.4 ml of reconstituted measles vaccine virus the other was left as a control,
with daily observation until 90% CPE was reached on the fifth day
post-inoculation. The tissue culture flask was then frozen and thawed three
times. The measles virus culture was then centrifuged at 3000 rpm for 20 min
using a cold centrifuge to obtain suspension of the viral particles which were
then dispensed in aliquots of 1 ml into cryovials, and stored at -20°C. The
polyethylene glycol (PEG) method of virus concentration was carried out
overnight at +4°C and then 5 μl of tween 80 detergents was added on ice and
vigorously shaken. 2 ml of ether were added and spun for 20 min at 3000 rpm in
a cold centrifuge. Purified MV HA Ag was collected from the interphase and
tested for hemagglutination activity as recommended by Rota et al. [12].
MV neutralization
assay
Serum was heat-inactivated at 56°C for 30
min. Tenfold serial dilution of the virus from10-1 to 10-5
(1 ml from the virus stock in 9 ml of EME medium), was prepared and 50 μl of
each dilution were distributed in 12 wells of 96 well micro titer plate except
those of the two last row (G-H) which was considered as positive control (vero
cells/medium), each row contained certain dilution (10-1 in the
first row A, 10-1.5, in row B, 10-2 in C and so on). Then
50 μl of 1:4 dilution of test serum (0.5 ml of serum in 1.5 ml of GEME medium)
was added to each virus dilution in duplicate the last two column (11-12)
represent negative control (virus dilutions/vero cells) then incubated at 37°C
for 1 h. Then 100 μl of vero cell line suspension were added and incubated at
37°C for 6 days with daily observation.
Neutralizing index
The neutralization index (NI) was calculated
according to Sperman and Karber method.
VNT antibody Index ≤1.5 was considered
protective.
Hemagglutination
assay (HA)
25 µl of PBS were added into 2 row of micro
titer plate and then 25 µl of virus suspension were added and mixed after that
25 µl from this well were transferred to the next well and mixed, this step
repeated to make twofold serial dilution along the row. 25 µl of 1% monkey red
blood cells were added to each well then incubated at 37°C for 1 h. The
reciprocal of the highest dilution that produce positive HA was considered as
the virus titer that would give one HAU.
Monkeys’ washed red
blood cells (RBCs)
2.5 ml of monkeys’ blood were collected by
venipuncture using 5 ml sterile disposable syringe containing 2.5 ml Alsevier
solution. The mixture was then dispensed in a tube and centrifuged at 1000 rpm
for 5 min then the supernatant was discarded. An equal volume to Alsevier
solution, of normal saline was added to the pelleted RBCs and the suspension
was centrifuged. This washing process was repeated three times and 1% suspension
of washed Monkey RBCs was prepared and preserved at 4°C.
Hemagglutination
inhibition test
The test was carried out using the MV HA Ag
to obtain I HA unit, from which the 4 HA units (standard working concentration)
was obtained. Then the HI was performed. HI antibody titers were measured by a
standard micro titer method using African green monkey red blood cells [13].
The HI test may be complicated by the
presence of non-specific inhibitors of viral hemagglutination and naturally
occurring agglutinins of the erythrocytes. Therefore, the sera were treated
before use. Serum was heat-inactivated at 56°C for 30 min. Non-specific
inhibitors of hemagglutination and non-specific agglutinins were removed from
test sera by the addition of 0.1 ml of each serum to 0.4 ml borate saline and
0.5 ml of a 25% suspension of acid washed kaolin. The mixture was spun at 1200
rpm for 10 min and 25 μl of monkey erythrocytes was added to remove
non-specific agglutinins, then spun at 1200 rpm for 10 min and supernate
collected in new labeled cryoviale [14]. 25 µl of PBS were added to each of 96
U-shaped wells of micro titer plate, then 25 µl treated sera were added to
well-1 and 25 µl from this well were transferred to well-2 and twofold serial
dilution was carried out along the row, this steps were done to each serum
sample, then 25 µl of 4HAU of antigen was added to each well of the serially
diluted sera except for the monkey erythrocytes control wells. The plate was
then allowed to stand for 1 h at 37°C. After that 25 µl of 1% monkey red blood
cells (baboon red blood cells), were added to each well and incubated for 1 h
at 37°C [15].
STATISTICAL ANALYSIS
Statistical analysis was carried out with
SPSS version 20.0 to test for equality of the means using the Chi square and T
test.
RESULTS
Description of the
study population
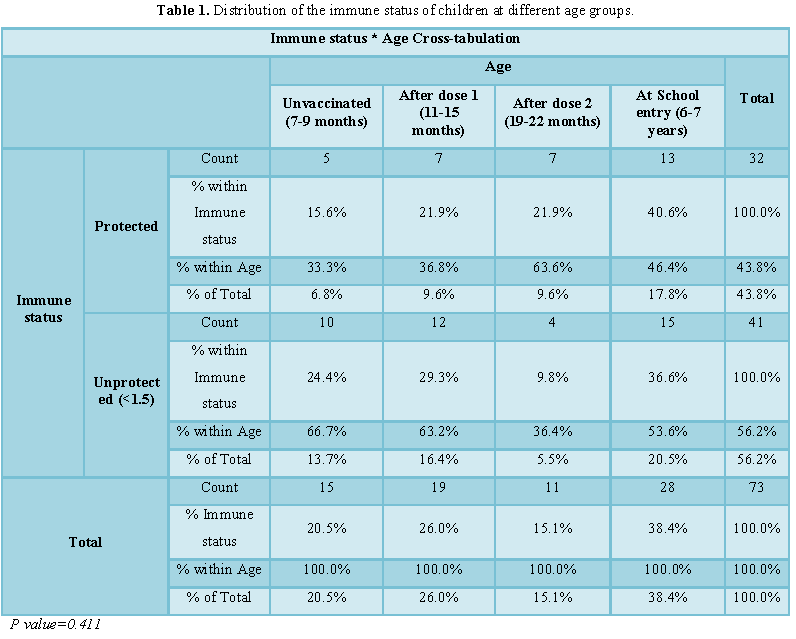
Overall 58 (79.5%) were vaccinated, 15
(20.5%) were unvaccinated. The age distributions of the infants recruited into
the study were as follow; the majority, 59.0% (28/73) were aged 6-7 years,
21.0% (19/73) were aged 10-15 months. The others 15/73 (14.0%) were aged 7-9
months and 6.0% (11/73) were aged 19-22 months (Table 1).
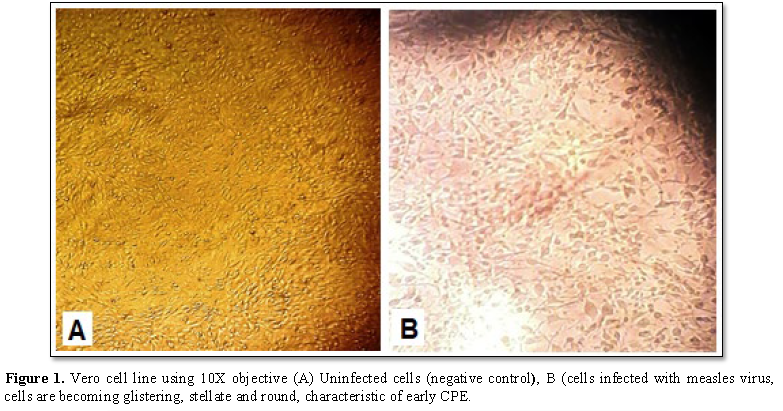
Cytopathic effect of
the measles virus in tissue culture
After 5 days, maximum development of the CPE
from measles virus vaccine in vero cells was seen using an inverted microscope.
Uninfected vero cells are shown in Figure
1A, are spindle cells. Infected cells (Figure
1B) are shining, round and stellate cells, followed by formation of
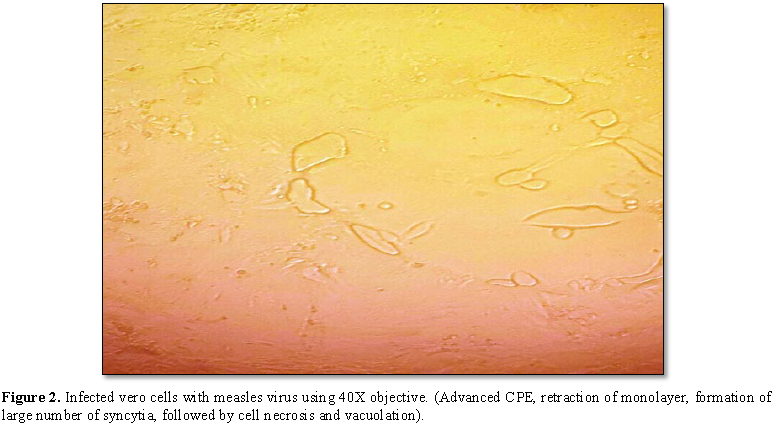
syncytia, (multinucleated cells) as well as which showed vacuolization of
cells, a CPE of special peculiarity (Figure
2).
Measurement of
antibodies against measles using neutralization test
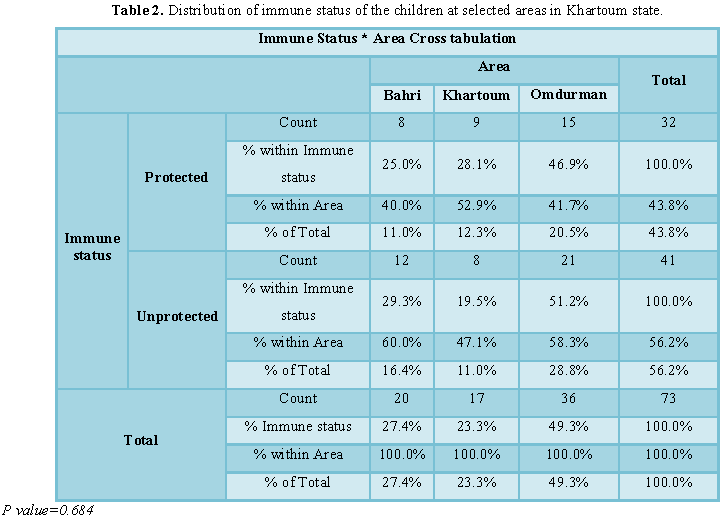
In this study 32 (43.8%) children of the population had a protective neutralizing antibody titer to measles virus (≥ 1.5) while 41 (56.2%) had detectable neutralizing antibody less than the protective titer. Before vaccination 5 (33.3%) of children were protected 10 (66.7%) unprotected, after the first dose 7 (36.8%) were protected and 12 (63.2%) were unprotected, after the second dose 7 (63.6%) were protected, 4 (36.4%) unprotected and at the School entry age 13 (46.4%) were protected, 15 (53.6%) unprotected (Table 1). There was no significant difference in the level of protection between the children in Bahri, Khartoum and Omdurman (p>0.05) as shown in Table 2.
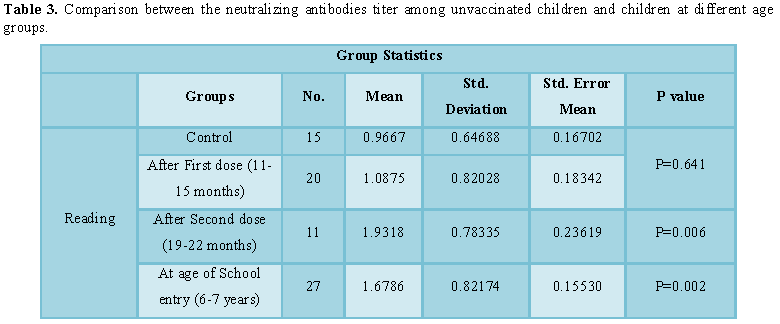
There was no significant difference between
titers of children who were not vaccinated and those who took the first dose of
vaccine p value=0.641. But there was a significant difference between children
of unvaccinated group and the other groups after the second dose and at age of
school entry, P=0.006, P=0.002, respectively (Table 3).
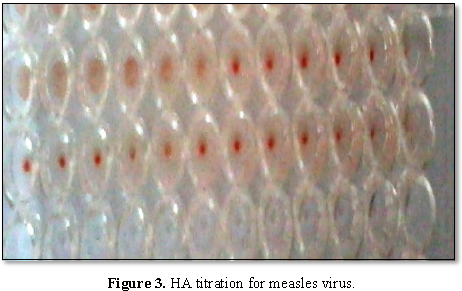
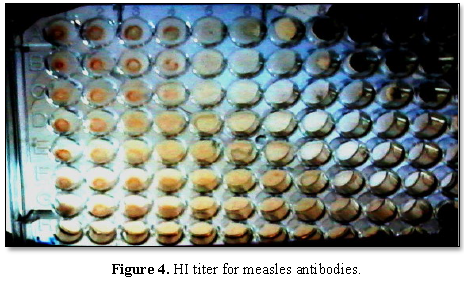
Measurement of antibodies against measles
using hemagglutination inhibition assay (HIA)
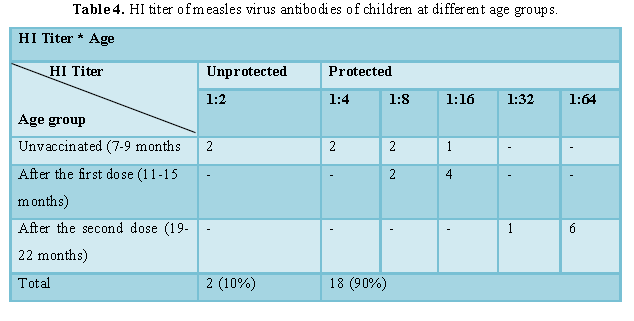
The agglutination of the RBCs by the concentrated virus appeared till the sixth well (1:64) which was considered as one HAU (64). Two wells back from the last well in which agglutination appeared, i.e., to wells at dilution (1:16) was considered as 4HAU (1 ml from concentrated virus in 15 ml PBS), as shown in Figures 3 and 4.
Complete inhibition of agglutination at 1:4 dilution
of serum was considered as the protection level (Titer=4) [19].
Out of the 20 children 18 (90%) had
protective antibodies level and 2 (10%) not protected.
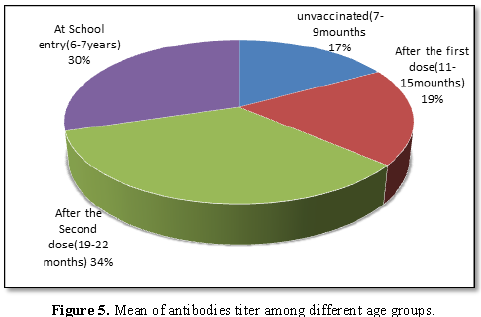
The titer of neutralizing antibodies rose slightly with advance in due to increased number of vaccinations (Table 4 and Figure 5).
DISCUSSION AND CONCLUSION
Antibodies to
the hemagglutinin (H) protein are the primary antibodies measured by
neutralization of virus infectivity in tissue culture [15], just like
hemagglutination inhibition (HI) test which is used to measure the antibodies
to the H-protein. Neutralizing antibodies play a very important role in
preventing re-infection; therefore, the neutralization test is used to evaluate
vaccine response and assess susceptibility to measles. This study examined the
level of measles hemagglutinating antibody in children in three selected
hospitals in Khartoum state. Using Neutralization test and Hemagglutination
inhibition (HI) test, which are classical techniques that have been employed in
prevalence studies and have good correlation with ELISA They reflect the level
of the population's antibodies [16]. The Hemagglutination inhibition (HI) test
is the most widely acceptable test in most developing countries. This is
because it is very sensitive and specific and easily performed in any laboratory
[17].
The result of
the neutralization test showed that (43.8%) had detectable neutralizing
antibodies to measles and (56.2%) had no detectable neutralizing antibodies,
i.e., are seronegative to the neutralization test, this could be due to vaccine
failure due to a break in the cold chain or malnutrition. Past studies have
shown that a break in the vaccine cold chain resulted in the inability to
develop measles protective antibodies [18].
Of particular
interest in this test is the relatively large number of vaccinated children in
all locations who were found not to be protected. This could be as a result of
vaccine failure; it could also be due to wrong information from parents about
the true vaccination status of their children.
Highest titer
was found in the age group at entry of school, this rising in titer may be due
to post vaccination infection which acted as booster dose. The 27 (46.6%)
unvaccinated children might be protected as a result of recovery from natural
infection or maternal antibodies in agreement with Adu et al. [19] in that
measles antibodies are found in children earlier than 9 months may be
maternally derived but not always. It is also possible that the antibodies were
acquired post exposure to circulating wild measles virus.
In this study
it is clear that the level or percentage of protection increase slightly after
the first dose and more after the second, this agrees with Stetler et al. [11].
who reported that more than 70% of children who received the first vaccination
before 10 months of age were shown to have neutralizing antibodies which lasted
at 15 months or older.
The
Hemagglutination inhibition (HI) test showed that out of the 20 children 18
(90%) had protective antibodies level (<4) and 2 (10%) not protected
(>4). This agreed with Onoja and Adeniji [20] in that out of the 175
children vaccinated in Ibadan, 60 (34.3%) had an antibody level not sufficient
to protect against measles infection and 115 (65.7%) were protected.
From
unvaccinated group 71.4% (5/7) showed a protective antibodies titer ≥ 4 this
result disagree with Omer et al. [21] in that children in rural Khartoum, had
detectable MV-antibodies at 5 months, which dropped sharply to nil at 8-9
months.
The antibody
titer rises according to the age of the children; this is in agreement with the
study of Onoja and Adeniji [20] who found that antibody level increases with
the age. There was no significant differences in antibodies titer of
unvaccinated children and after the first dose of vaccine p=0.641, which may be
due interference of maternal antibodies with the vaccine, this agrees with
Ibrahim et al. [8] in that because the interference of maternal antibodies the
first measles vaccination is carried out between the ages of 12 and 15 months
in most industrialized countries. But a significant difference appeared after
the second dose p=0.002 and at age of school entry p=0.006 .This means a
booster dose is needed in order to attain protection against measles infection.
This agrees with Masae et al. [8], in that protective immunity induced by
vaccination may not be life long without being boosted either via a second dose
of vaccine or natural infection.
From the
comparison between the two test used, HI was found to be more sensitive in
evaluating antibodies titer and that may be as a result of treatment of the
virus stock using PEG/Tween 80 treatment, which increased the potency of virus
H Ag. This is in agreement with Rota et al. [12] in that the polyethylene
glycol (PEG) method of virus concentration increased the potency of virus
hemagglutinin antigen. Also this agrees with Motayo et al. [17], in that
hemagglutination inhibition (HI) test is the most widely acceptable test in
most developing countries .This is because it is very sensitive and specific
and easily performed in any laboratory.
Finally these
results allow us to conclude that the titer of measles antibodies at different
age groups of children in Khartoum State, increased with age and was related to
the number of vaccinations, out of 14 serum samples screened using HI and
NT100% (14/14), 35.7% (5/14) were protected, respectively. This is evidence
that HI could be a standard test in laboratory for measles diagnosis. Due to
the importance of the disease and the lack of definitive treatment to it,
procedures for evaluating the efficacy of vaccine must be maintained and
further studies are required.
1. Kalter
SS, Herberling RL, Barry JD (1991) Detection and titration of measles virus
antibody by hemagglutination inhibition and by dot immunobinding. J Clin Microbiol
29: 202-204.
2. Olivé
JM, Aylward RB, Melgaard B (1997) Disease eradication as a public health
strategy: Is measles next? World Health Stat Q 50: 185-187.
3. Youwang
Y, Ping W, Feng C (2001) Serological and epidemiological effects and influence
factors of primary immunizations with current live attenuated measles vaccine
(Hu 191) among infants aged 6-15 months. Vaccine 19: 1998.
4. Clements
CJ, Cutts FT (1995) The epidemiology of measles: Thirty years of vaccination.
Curr Top Microbiol Immunol 191: 13-33.
5. Argüelles
MH, Orellana ML, Castello AA, Villegas GA, Masini M, et al. (2006) Measles
virus-specific antibody levels in individuals in Argentina who received a
one-dose vaccine. J Clin Microbiol 44: 2733-2738.
6. Mossong
J, O'Callaghan CJ, Ratnam S (2000) Modeling antibody response to measles
vaccine and subsequent waning of immunity in a low exposure population. Vaccine
19: 523-529.
7. Kumar
ML, Johnson CE, Chui LW, Whitwell JK, Staehle B, et al. (1998) Immune response
to measles vaccine in 6 month old infants of measles seronegative mothers.
Vaccine 16: 2047-2051.
8. Ibrahim
SA, Abdallah A, Saleh EA, Osterhaus AD, De Swart RL (2006) Measles
virus-specific antibody levels in Sudanese infants: A prospective study using
filter-paper blood samples. Epidemiol Infect 134: 79-85.
9. Itoh
M, Okuno Y, Hotta H (2002) Comparative analysis of titers of antibody against
measles virus in sera of vaccinated and naturally infected Japanese individuals
of different age groups. J Clin Microbiol 40: 1733-1738.
10. Mitchell
LA, Tingle AJ, Décarie D, Lajeunesse C (1998) Serologic responses to measles,
mumps and rubella (MMR) vaccine in healthy infants: Failure to respond to
measles and mumps components may influence decisions on timing of the second
dose of MMR. Can J Public Health 89: 325-328.
11. Stetler
HC, Orenstein WA, Bernier RH, Herrmann KL, Sirotkin B, et al. (1986) Impact of
revaccinating children who initially received measles vaccine before 10 months
of age. Pediatrics 77: 471-476.
12. Rota
JS, Hummel KB, Rota PA, Bellini WJ (1992) Genetic variability of the
glycoprotein genes of current wild-type measles isolates. Virology 188:
135-142.
13. Orvell
C (1976) Identification of paramyxovirus-specific hemolysis inhibiting
antibodies separate from hemagglutinating-inhibiting and
neuraminidase-inhibiting antibodies. Acta Pathol Microbiol Scand B 84: 441-450.
14. Gershon
AA, Krugman S, Lennette EH, Schmidt NJ (1979) Diagnostic procedures for viral,
rickettsial and chlamydial infections. Am Public Health Assoc, Washington DC,
pp: 665-693.
15. Norrby
E, Gollmar Y (1975) Identification of measles virus-specific
hemolysis-inhibition antibodies. Infect Immun 11: 231-239.
16. Cox
MJ, Azevedo RS, Massad E, Fooks AR, Nokes DJ (1998) Measles antibody levels in
a vaccinated population in Brazil. Trans R Soc Trop Med Hyg 92: 227-230.
17. Motayo
BO, Okonko IO, Faneye A, Adu FD (2011) Preparation of high potent measles
hemagglutinating antigen from wild type measles virus in Ibadan, Nigeria. N Y
Sci J 4.
18. Adu
FD, Ikusika A, Omotade O (1997) Measles outbreak in Ibadan: clinical,
serological and virological identification of affected children in selected
hospitals. J Infect 3: 241-245.
19. Adu
FD, Odoemele FC, Bamgboye E (1999) Effect of measles antibodies in the breast
milk and sera of mother on seroconversion to measles vaccine. Afr J Biomed Res
2: 7-11.
20. Onoja
AB, Adeniji AJ (2013) Kinetics of measles antibody by hemagglutination
inhibition assay in children in south-west and north-central Nigeria. Int J
Infect Dis 17: e552-e555.
21. Omer
MI, el dawla IG, Nicolas A, Roumiantzeff R, Lapeyssonie L (1986) Humoral
antibody responses to biannual multiantigen vaccination: Report of a field
trial on children in Sudan. Ann Trop Pediatr 6: 243-248.
-
 Table 1
Table 1 -
Table 2
-
Table 3
-
Table 4