
13129
Views & Citations12129
Likes & Shares
The cultivation and conservation of common bean is faced with various phytosanitary problems related to biotic constraints such as fungi, insect pests and rodents on the one hand and abiotic constraints such as temperature and humidity on the other hand [7]. Fungal attacks on P. vulgaris L. crops cause considerable production losses and damage in humid tropical biotopes. In Cameroon, the species Sclerotinia sclerotiorum is the main cause of white rot of the common bean crop in the field. This pathogen causes white rot in about 408 plant species (including P. vulagaris L.) of about 275 different varieties [8]. Despite much work to control this pathogen, it continues to be a major cause of yield loss. The use of synthetic fungicides is one of the most effective methods of controlling this type of pathogen [9]. However, good quality synthetic products are rare and expensive in rural areas they very often have a harmful residual effect on human health and the biocenosis; they disturb ecological balances and pollute the environment [10-12].
Currently, various studies focus on the use of plants with fungicidal and insecticidal properties against diseases and insect pests of crops [13-15]. To this end, several works have shown and continue to show the antifungal potentialities of plant essential oils on the in vitro and in vivo growth of some fungi. Notably, the work of Yilar [16] on the in vitro inhibition of the growth of fungi such as Rhizoctonia solani J.G. Kuhn, S. sclerotiorum (Lib.) de Bary by the essential oils (EOs) of Vitex agnus-castus L. and Myrtus communis L; The use of Eucalyptus storigeriana EO on Alternaria solani [17] and the use of Commiphora molmol EO on the reduction of mycelial growth of Aspergillus flavus, Fusarium oxysporum and A. alternata [18]. The objective of this work is to control the mycelial growth of three strains of S. sclerotiorum fungi with the EO of two plants, namely T. vulgaris and C. Citratrus.
MATERIAL AND METHODS
Material
Plant material
For the present work, two species of plants are used, namely thyme (T. vulgaris) and lemongrass (C. citratrus) (Figure 1). The leaves of T. vulgaris were purchased dry at the local medicinal plant market (Dakar market, Douala-Cameroon) while the leaves of C. citratus were harvested manually in the field and dried for 2 weeks in the air and out of the sun.
Chemical Material
The chemical material used is Banko-Plus 650 SC (Chlorothalonil at 550 g/L + Carbendazim at 100 g/L).
Fungal material
The fungal material consisted of two strains of fungi from the Central Region, town of Biakoa (Bia 1 and Bia 2) and one strain from the Littoral Region, town of Njombé (Njo 2).
Methods
Extraction yield of essential oils
The extraction yields of lemongrass (HEC) and thyme (HET) essential oils were calculated in relation to the weights of plant material according to the following formula: [19].
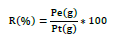
Where R: Yield; Pe: Weight of the extract (essential oil) Pt: Total weight of dried plant material
In vitro evaluation of the effect of essential oils on the growth of different strains of S. sclerotiorum
Preparation of the culture medium
The preparation of the culture medium was done by taking 65 g of agar (Sabouraud Agar) [20] which are introduced into 1000 ml of sterile distilled water. The mixture is heated on a hot plate equipped with a magnetic stirrer for 10 min to homogenize the solution. The resulting solution is then autoclaved for 45min at 120°C and stored in the refrigerator for future use.
Isolation and purification of pathogen strains
Pure strains were isolated from infected bean leaves showing symptoms of the disease. The leaves were harvested in the field and then taken to the laboratory where they were washed several times with tap water, disinfected with a cotton soaked in 95% alcohol and then flame dried. The infected parts of the leaf (bearing the necroses) were removed using a scalpel and incubated in Petri dishes containing Sabouraud Agar medium. The Petri dish was then sealed with food film and incubated in a culture room at 22-24°C under a 12h photoperiod. Mycelia grew from the explant and reached sufficient growth after 5 days, ready for purification. Purification was carried out by successive subcultures of a fragment taken from the growth front of mycelium on Sabouraud Agar medium. This operation was repeated several times until pure cultures were obtained that will be kept in pill containers, containing sterile distilled water [21,22]. Identification of the pathogen S. sclerotiorum was done through the observation of conidia on the microscope and an identification key [23,24].
Essential oil extraction by hydrodistillation
The extraction of essential oils was carried out by hydro distillation using a hydro distillation apparatus type Clevenger [25]. The operation, which lasts 3 h, consists in introducing 100 g of dried plant material (Thyme and Lemon grass) in a glass flask, then submerged the plants with a quantity of distilled water (1L). Using a heating flask, the mixture is brought to a boil. The water vapors carry away the volatile essential oils, which are then condensed by a cold condenser (cooling coil) that transforms them into liquid. The essential oil is lighter than water and floats to the surface. It is recovered, measured and weighed to determine the yield. The HET and HEC were dehydrated with sodium sulfate (Na2SO4) and kept in a refrigerator at 4°C and in the dark until analysis and future use [26,27].
Determination of the different doses of essential oils
To obtain the different doses of HEC and HET, volumes of 0.23, 0.45 and 0.9 ml were taken with a pipette and then, introduced respectively in 29.77, 29.55 and 29.1 ml of nutrient media cooled to 45 °C in order to obtain Sabouraud Agar media with respective concentrations of essential oils of 0.75, 1.5 and 3 ml/ml. For the synthetic fungicide, a stock solution was previously prepared by diluting 1 ml of Banko-plus 650 SC (550 g/L chlorothalonil + 100 g/L Carbendazim) in 200 ml of sterile distilled water. From this stock solution, a volume of 0.2 mL was pipetted into 29.8 mL of Sabouraud Agar nutrient medium. These culture media were poured into 90 mm Petri dishes. Agar discs of 6 mm diameter, bearing the mycelium of each isolate, were individually placed in the Petri dishes in the center of the different solidified culture media. The susceptibility of the fungus was assessed by measuring the radial growth of the mycelium every 24 h after two days of inoculation. The experiment lasted 08 days and was repeated 3 times.
Radial growth
The average growth (D) of S. sclerotiorum mycelium is determined by daily measurements of necrosis 48 h after incubation using a graduated scale. Each diameter is respectively measured on one of two lines forming a right angle through the center of the seeded explant (Figure 2). The percentages of inhibition (I%) of the different doses of EO and of the positive control are calculated after 7 to 9 days of fungal growth.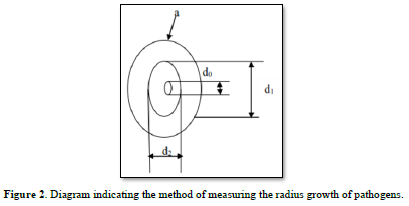
Mean radial growth and percent inhibition are calculated using the following formulas: [28].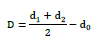
Where do: Diameter of the explant; d1 and d2: Perpendicular diameters of the pathogen; D: Mean radial diameter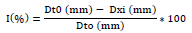
Where I (%): Percent inhibition; Dt0: Average diameter of necrosis in the negative control; Dxi: Average diameter of necrosis in the HE or positive control
Evaluation of fungicidal or fungistatic activities of essential oils
After incubation, the treatments in which the growth of the fungus was completely inhibited are counted and the explants are transferred to a new culture medium without EO. The extract is described as fungistatic if mycelial growth restarted and as fungicidal if not [29,30].
Determination of minimum inhibitory concentrations
The values of the different inhibition percentages were used to determine the minimum inhibitory concentrations from the formula of [28]. From the linear regression equation between the neperian logarithms of the concentrations on the x-axis and the growth inhibition percentages on the y-axis, the concentrations reducing growth by 50 and 90% are determined [31].
Correlations between concentration and inhibition
The correlation between concentration and percentage inhibition was determined from the equation y = ax + b with x = concentration, and y = percentage inhibition. In this case, if a < 0, the slope is negative; if a > 0, then the slope is positive; if r is between 0.8 and 1, then the correlation is perfect and positive; if r is between -0.8 and -1 then the correlation is perfect and negative; if r < 0.8 then the correlation is positive but imperfect; if r > -0.8 then the correlation is negative but imperfect [32].
Data analysis
Data used were subjected to one-way analysis of variance (ANOVA) using SPSS 20.0 software. The multiple comparisons of means were performed using the Last significant difference (LSD) test at P ˂ 0.05 to separate them when the analysis of variance was significant. The graphs were presented and the average grouping tables were produced in the Excel 2013 spreadsheet.
RESULTS
Extraction yield of essential oils
The two extractions having been made by hydrodistillation, yields of 1.5 ± 0.25% and 0.5 ± 0.1% were obtained with C. citratus and T. vuglaris respectively (Table 1).
Effect of essential oils on the growth of different strains of S. sclerotiorum
Virulence of the strains used
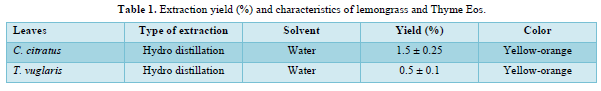
The data in Table 2 show the growth rate of the three strains of S. sclerotiorum used in this work regardless of the extracts and concentrations used. It appears that the Njo 2 and Bia 1 strains are the most virulent during 8 days of culture with respective radial growth diameters of 7.12 and 5.95 cm against 4.88 cm for the Bia 2 strain which thus proves to be the least virulent during the 8 days of culture.
Effects of lemongrass essential oils (HEC) on the growth of different strains
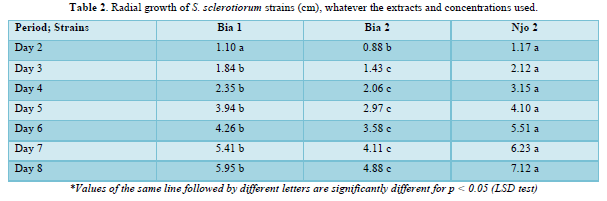
The results on the effect of HEC on the radial growth of the different strains of S. sclerotiorum tested show a considerable inhibition of the said radial growth at the different concentrations. A significant difference was observed between these 3 concentrations (C1, C2 and C3) which show different behaviors at the 5% threshold according to the LSD test. However, the Njo 2 strain proved to be more resistant to the different concentrations of HEC because, after 8 days of treatment, a high mycelial growth was observed at concentrations C1= 0.75ml/ml and C2= 1.5 ml/ml, i.e.7.12 and 7.07 cm respectively; these results are not significantly different from the one observed in the control treatment (T-= 0ml/ml) which showed a growth of 7.3 cm at the same time. On the other hand, the concentration C3 =3ml/ml showed a considerable inhibition (3.75 cm) which is statistically different from the previous three (Figure 3). The results also show in general that the strains of the Bia 1 and Bia 2 Center have almost the same behavior towards HEC.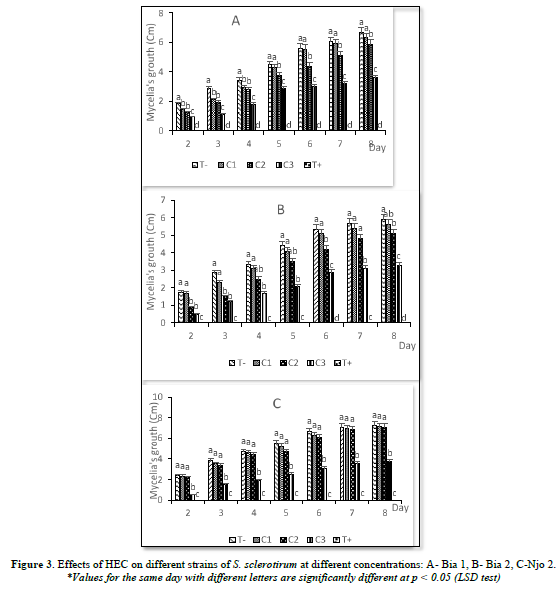
Effect of essential oils of thyme (HET) on the growth of strains
The HET presented a very high antifungal property with, in most cases, a percentage of inhibition of the order of 100%. A total inhibition of the growth of Bia 1 and Bia 2 strains and this at all concentrations (C1, C2 and C3) was observed. The Njo 2 strain showed a slight resistance to HET at concentrations C1= 0.75 ml/ml and C2= 1.5 ml/ml, with radial growths of 7.12 and 6.5 cm respectively after 8 days of treatment. This strain, on the other hand, underwent a total inhibition of its growth at the concentration C3= 3ml/ml (Figure 4).
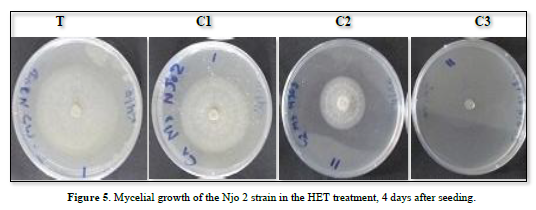
The effect of HET evaluated on the Littoral strain (Njo 2), showed a significant repression of the fungus development. Compared to the control (0 ml/ml), the different concentrations used (C1: 0.75 ml/ml; C2: 1.5 ml/ml and C3: 3 ml/ml) showed significant inhibition of mycelial growth of the S. sclerotirum strain at each observation period. The inhibition here is proportional to the different concentrations, as it increases with EO concentration (Figure 5).
The results of the inhibition percentages obtained after 5 days of treatments show that the different essential oils significantly inhibited the mycelial growth of the strains at the different concentrations tested. A significant difference on the efficacy of the different concentrations within the same EO was observed for the three tested strains. As an illustration, a significant difference was observed between the percentages of inhibition obtained at the concentrations C1, C2 and C3 tested on the Njo 2 strain with the EO; we have respectively 9.82; 20.63 and 54.18 % (Table 3). Moreover, between the two HE, the results obtained show that HET are more effective than HEC. A significant difference is observed between these two EO when comparing the behavior of the same concentration on the same strain. Taking as an example the Bia 1 strain, a significant difference at the different concentrations (C1, C2 and C3) is observed: HEC (3.57; 14.55 and 16.29%) versus HET (100; 100 and 100%) respectively (Table 3).
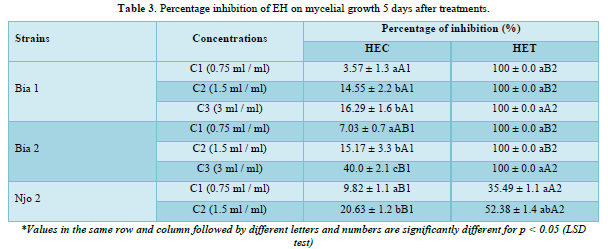
The triplet (a, b and c) was used to compare in the same EO and for the same strain, the behavior of the different concentrations; the triplet (A, B, C) was used to compare in the same EO and for the different strains, the behavior of the same concentration and, the doublet (1, 2) was used to compare between the different EO and for the same strain, the behavior of the same concentration.
Fungicidal/fungistatic properties of extracts
For the fungicidal/fungistatic test conducted on the different strains of S. sclerotiorum, only the concentrations of essential oils that completely inhibited mycelial growth are mentioned. The HECs, at all concentrations tested did not totally inhibit mycelial growth; therefore, this test could not be performed for these extracts. With the HETs, a fungistatic activity was observed at concentration C1= 0.75 ml / ml in the Bia 1 and Bia 2 strains, while concentrations C2= 1.5 ml / ml and C3= 3 ml / ml presented a fungicidal activity on these same strains. Njo 2 strain had complete inhibition of mycelial growth only at its C3 concentration for fungicidal activity with HET (Table 4).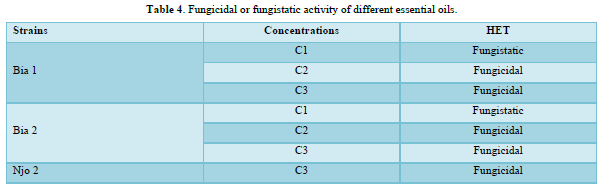
Correlation tests between concentrations and inhibition percentages
The purpose of this test was to see if there is a linear relationship between the increase in mycelial growth inhibition of the pathogen and the different concentrations of the essential oils. The regression equations obtained with HEC showed increasing linear relationships between concentrations and inhibition percentages. All strains tested, showed regression lines with positive slopes: y= 20.841x - 7.2933; y= 7.4058x + 72.747 and y= 23.594x + 13.173, respectively for the strains Bia 1, Bia 2, and Njo 2 (Figure 6). For these three strains, the correlation coefficient values: r= +0.93, r= +0.87, and r= +0.87 were obtained respectively (Table 5).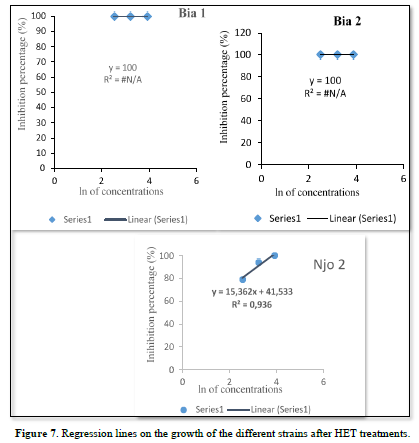

Minimum inhibitory concentrations MIC50 and MIC90 on some strains used
From the regression lines obtained after the correlation tests, the concentrations inhibiting by 50% (MIC50) and 90% (MIC90) the growth of the different pathogen strains were determined.
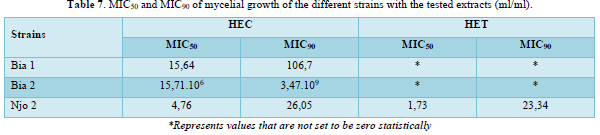
The lowest inhibitory concentrations were obtained with HET, justifying their greater efficacy compared to HEC. The MIC50 obtained with HEC on all three strains tested ranged from 4.76 ml/ml for the Njo 2 strain to 15.71.106 ml/ml for the Bia 2 strain. The MIC90 vary from 20.05 m/ml in the Njo 2 strain to 3.47.109 ml/ml in the Bia 2 strain for the same treatment. On the other hand, for the treatment of these same three strains with HET, the MIC50 and MIC90 could only be defined for the Njo 2 strain where they were respectively 1.73 ml/ml and 23.34 ml/ml. On the Bia 1 and Bia 2 strains, growth inhibition was total and therefore it was not possible to determine them. (Table 7).
DISCUSSION
The yields obtained after extraction of the essential oils of the two investigated plants (1.5% for HEC and 0.5% for HET) show that this parameter varies from one plant species to another. The yield of HEC obtained in this study is higher than those obtained by François et al. (2009) and which are respectively 0.67 and 0.7% but, lower than those obtained on the same plant species by Kanko [33], Konfo [34] and Kpatinvoh [35] and which are around 4.31%. The difference in yield of these two plants can be explained by the influence of some extrinsic factors such as the geographical origin of the raw material; the cultural conditions such as sowing date, harvesting date, phytosanitary treatments, fertilizer use, and harvesting techniques [36,37]. Concerning HET, the results of the extraction yield agree with those obtained on the same plant by Rachid [38] who obtained a yield of 0.65%.
Generally speaking, the results of this work show a significantly positive effect of essential oils, especially HET, in the various tests carried out on white rot of beans in the laboratory. The different strains tested showed different behaviors from each other. The Littoral strain (Njo 2) was the most aggressive (most virulent) with a necrosis growth of 7.2 cm at day 8. Such results were obtained by Ngoh Dooh [39] and Nchare [40] in studies based on the characterization of some fungal strains from the Central, Southern and Littoral regions.
The fact of obtaining strains more resistant (Njo 2) to EO than others would be due to the non-specificity that these different strains would present at the membrane level. Indeed, antifungals can be contact (the antifungal acts at the membrane of the fungus) or systemic: the antifungal acts inside the cell [41]. In either case, specific membrane or intracellular receptors may be essential for the expression of the antifungal's biological activity [42].
HET and HEC significantly inhibited the growth of the different fungal strains tested. However, inhibition percentages of the order of 100% were obtained with HET at all concentrations on the strains Bia 1 and Bia 2 and, at the concentration C3= 3 ml/ml on the strain Njo 1. The work of Kpatinvoh [35] showed the effectiveness of some essential oils including those of C. citratus L. of C. nardus L. and Eucalyptus saligna on the weathering mycoflora of Vignia unguiculata L. in South-Benin. Similarly, Sirima [43] showed the efficacy of C. schoenanthus EO with inhibitions ranging from 26 to 100% at doses of 1%, 5%, 10%, 50% and 100% in vitro conditions.
These results obtained showing considerable inhibition of mycelial growth (100%) at different concentrations of EO are in line with those of Rachid [38] who, comparing in their work the antifungal activity of three essential oils (T. vulgaris, C. limonum and Mentha spicata), on the growth of some dermatophytes had a significant effect with the essential oils of T. vulgaris at concentrations greater than or equal to 2% against equally significant effects but, at high concentrations of 4 and10% respectively for the essential oils of M. spicata and C. limonum. According to [44], the antifungal efficacy of EO is related to an active substance called thymol.
The MIC50 and MIC90 of the different EO were determined with the different strains tested. An important variability was observed; the HET presenting the lowest values, which justifies their effectiveness and consequently their fungicidal potential. This variability could be explained by the fact that phytopathogenic fungi do not react in the same way to biopesticides. Such results were obtained by Carlton [45] who showed that phytopathogenic fungi act differently in the presence of biopesticides. These results highlighting the higher efficacy of HET via low MIC (50 and 90) (1.73 and 23.34 ml/ml) compared to HEC: (4.76 and 26.05 ml/ml) respectively, are in line with those obtained by Kpatinvoh [35] who obtained high inhibition percentages on cowpea spoilage mycoflora with MIC values between 0.05 and 1 µl/ml. In addition, Mboussi [46] also obtained similar results on the radial growth of some fungi with low MIC 50, 90 and 95 of the aqueous extracts of Thevetia peruviana, i.e 2.9; 4.3 and 4.48 ml/ml.
The effectiveness of Banko-Plus 650 SC has been demonstrated once again by completely inhibiting the mycelial growth of the different strains of S. sclerotiorum tested. This product is known to have a direct and toxic action against some fungi in general and S. sclerotiorum in particular through a persistence of its active principles, namely Chlorothalonil and Carbendazim.
CONCLUSION
The objective of this work was to evaluate the antifungal activity of essential oils of two aromatic plants against three strains of S. sclerotiorum responsible for white rot of bean. The different essential oils used showed significantly high efficacy on the reduction of mycelial growth with a particularly satisfactory behavior obtained with HET (100% inhibition) at all concentrations. At different concentrations and according to the two EO used, the Njo 2 strain proved to be the most aggressive while the Bia 2 strain was more sensitive to the different treatments. These different extracts could be, depending on the efficiencies presented, exploited as biodegradable antifungal substances able to control white rot of beans.
- Wortmann CS (2006) Phaseolus vulgaris (common bean): Plant Ressources of Tropical Africa/Ressources Végétales de l’Afrique tropicale (Prota 1) CIAT: Cali, Colombia.
- Anonyme (2005) Application de la biotechnologie à la lutte contre les maladies du haricot. N° 27; Le CIAT en Afrique.
- Pabra (2006) Nutrition et santé: la vision de PABRA. Alliance Panafricaine de Recherche sur le Haricot 3: 2.
- Wortmann CS, Kirkby RA, Elude CA, Allen DJ (1998) Atlas of Common Bean (Phaseolus vulgaris ) Production in Africa (Prota 2). CIAT: Cali, Colombia.
- Anonyme (2010) Annuaire des Statistiques du Secteur Agricol du Cameroun. Available online at: minader.cm/documents-destatistiques-de-la-direction-des-études-etstatistiques-agricoles
- Pamo TE, Boukila B, Tankou MC, Tendonkeng F, Kana J-R, et al. (2005) Effet de différentes sources d’azote sur la croissance et le rendement du haricot commun (Phaseolus vulgaris) à l’Ouest Cameroun. Cameroon J Exp Biol 01(01): 1-7.
- Diby YKS, Tahiri YA, Akpesse AAM, Tra Bi CS (2015) Graine: Haricot. In: Agriculture en Afrique Tropical, Bruxelles. DGCI. pp: 337-355.
- Boland GJ, Hall R (1994) Index of plant hosts of Sclerotinia sclerotiorum. Can J Plant Pathol 16: 93-108.
- Haubruge E, Shiffers B, Gabriel E. et Verstraeten C, (1988) Etude de la relation dose-efficacité de six insecticides à l’égard de Sitophilus granarius L., S. oryzae L., S. zeamais Mots. (Coleoptera, Curculionidae). Mededelingen Faculteit Landbouwwetenschappen Rijksuniversiteit Gent. 53/2b, 719-726.
- Condor-Golec AF (2007) Effect of neem (Azadirachta indica A. Juss) insecticides on parasitoids. Rev Per Biol 14(1): 69-74.
- Regnault-Roger C (2007) Biopesticides: une alternative aux pesticides conventionnels? Mer, Littoral, Lacs et Cours d’eau 73: 60-64.
- Dohou N, Yamni K, Badoc A. et Douira A (2004) Activité antifongique d’extraits de Thymelaea lythroides sur trois champignons pathogènes du riz. Bull Soc Pharm Bord 143: 31-38.
- Doumbouya M, Abo K, Lepengue AN, Camara B, Kanko K, et al. (2012) Comparing in vitro activities of two synthetic fungicides and two essential oils on soil fungi of vegetable crops in Côte d’Ivoire. J Appl Biosci pp: 1-13.
- Messaoundene H, Mouhou N (2017) Etude de la toxicité des huiles essentielles contre les ravageurs des denrées stockées. Etude de la toxicité des huiles essentielles contre les ravageurs des denrées stockées. Mémoire de Doctorat, Faculté des Sciences de la Nature et la Vie, Université Abderrahmane MIR-Bejaia, Algérie pp: 35.
- Lipi P, Agindotan BO, Burrows ME (2021) Antifungal activity of Plant-Derived Essantial Oils on Pathogens of Pulse Crops. Plant Dis 105: 1692-1701.
- Yilar M, Bayan Y, Onaran A (2016) Chemical composition and antifungal effects of vitex agnus-castus and myrtus communis L. plants. Notulae Botanicae Horti Agrobotanici Cluj Napoca 44(2): 466-471.
- Tomazini EZ, Pauletti GF, Ribeiro RTS, Moura S, Schwambach J (2017) In vitro and in vivo activity of essential oils extracted from Eucalyptus staigeriana, Eucalyptus globulus and Cinnamomum camphora against Alternaria solani Sorauer causing early blight in tomato. Scientia Horticulturae 223: 72-77.
- Perveen K, Bokhari NA, Siddique I, AlRashid SAI (2018) Antifungal Activity of Essential Oil of Commiphoramolmol Oleo Gum Resin. J Essent Oil-Bear Plants 21: 667-673.
- Ngoh-Dooh JP, Ambang Z, Tih-Ewola A, Heu A, Kosma P, et al. (2014a) Screening and the effect of extracts of Thevetia peruviana on the development of colletotrichum gloeosporioides, causal agent of cassava anthracnose disease. E3 J Agric Res Develop 4(4): 54-65.
- Sabouraud R (1892) Contribution a l’etude de la trichophytie humaine. Etude clinique, microscopique et bacterioloqique sur la pluralité des trichophytons de l’homme. Ann Dermatol Syphilol 3: 1061.
- Ondo AS (2006) Charactérisation de quelques isolats de megakarya agent causal de la pourriture brune des cabosses de cacaoyer (Theobroma cacao L) Mémoire de DEA, Université de Yaoundé I pp: 58.
- Nyassé S (1992) Structure d’une population de Phytophthora spp. des cacaoyères camerounaises atteintes de pourriture brune. Mémoire de diplôme de Recherche Universitaire ENSAT, Toulouse pp: 43.
- Vaz PDC (1987) IMP Description of Fungi and Bacteria. 92, sheet 916.
- Hsieh WH, Goh TK (1990) Cercospora and similar fungi from Taiwan. Maw Chang Book Compagny, Taiwan. Available online at: bcrc.firdi.org.tw/fungi/fungal
- Oussou KR (2009) Etude chimique et activités biologiques des huiles essentielles de sept plantes aromatiques de la pharmacopée ivoirienne. Thèse de Doctorat Unique. Laboratoire de Chimie Organique et Structurale, UFR SSMT, Université de Cocody-Abidjan pp: 241.
- Bajpai VK, Shukla S, Kang SC (2008) Chemical composition and antifungal activity of essential oil and various extract of Silene armeria Bioresour Technol 99: 8903-8908.
- Willem JP (2004) Les huiles essentielles, Médecine d’avenir. Ed, Dauphin, Paris pp: 311.
- Singh G, Padvay RK, Narayanam CS, Padmhurmeri KP, Rao GP (1993) Chimical and fungistatic investigation out the essential oil citrus. Pers. Z. den zee Hal fur pflan und flanz 100: 69-74.
- Kishore N, Mishra AK, Cham Souria JPN (1993) Fungitoxicity of essential oils against dermatophytes. Myc 36: 211-215.
- Pandey DK, Chandra H, Tripathi NN (1982) Volatile fungitoxicity Activity in higher plants special reference to that of Callistemun lanceolatus J Phytopathol 105: 175-182.
- Dohou N, Yamni K, Badoc A, Douira A (2004) Activité antifongique d’extraits de Thymelaea lythroides sur trois champignons pathogènes du riz. Bull Soc Pharm Bord 143: 31-38.
- Heu A (2013) Effet des extraits de graines de Thevetia peruviana sur la germination des spores de Phytophthora megakarya et de Colletotrichum gloeosporioides in vitro. Mémoire de Master. Université de Yaoundé I pp: 52.
- Kanko C (2010) Contribution à l’étude phytochimique de plantes médicales et aromatiques de Côte d’Ivoire. Activité analgésique et antiinflammatoire de stérols isolés de l’écorce de Parkia biglobosa. Thèse de doctorat, Université de Cocody-Abidjan., Abidjan. pp: 224.
- Konfo C, Ahoussi-Dahouenon E, Sessou P, Yehouenou B, Djenontin S, et al. (2012) Stabilization of Local Drink "Tchakpalo" produced in Benin by addition of Essential Oil Extracted from Fresh leaves of Cymbopogon citratus. Int Res J Biol Sci 1(8): 40-49.
- Kpatinvoh B. Adjou Euloge S, Dahouenon-Ahoussi E, Konfo TRC, Atrevi B, et al. (2017) Efficacité des huiles essentielles de trois plantes aromatiques contre la mycoflore d’altération du niébé collecté dans les magasins de vente du Sud-Bénin. J Appl Biosci 109: 10680-10687.
- Benini C (2007) Contribution à l’étude de la diversification de la production d’huiles essentielles aux Comores. Mémoire d’ingéniorat. Université Gembloux, pp: 109.
- Aprotosoaie AC, Spac AD, Hancianu M, Miron A, Tanasescu VF, et al. (2010) The chemical profile of essential oils obtained from fennel fruits (Foeniculum vulgare Mill.). Farmacia 58(1): 46-54.
- Rachid I, Abdeslam L, Khadija M (2014) Study of the antifungal activity of essential oils of three moroccan aromatic plants. Int J Innov Sci Res 12: 499-505.
- Ngoh-Dooh JP, Ambang Z, Ndongo B, Heu A, Kuate TW (2014b) Effect of extracts of Thevetia peruviana (Pers.) K. Schum on development of Phytophthora megakarya causal agent of black pod disease of cocoa. J Appl Biosci 77: 6564-6574.
- Nchare SF (2013) Évaluation du potentiel antifongique des extraits de graines de Jatropha curcas sur le développement in vitro des souches de Phytophthora megakarya. Mémoire de Master. Université de Yaoundé I pp: 52.
- Zacchino S, Rodriguez G, Santecchia C, Pezzenati G, Giannini F, et al. (1998) In vitro studies on mode of action of antifungal 804’-neolignans occuring in certain species of Virola and related genera of Myristicaceae. J Ethnopharmacol 62 (1): 35-41.
- Mboussi S, Ambang Z, Ndogho P, NgohDooh J, Manga Essouma F (2016) In vitro antifungal potential of aqueous seeds extracts of Azadirachta indica and Thevetia peruviana against Phytophthora megakarya in Cameroon. J Appl Life Sci Int 4(4): 1-12.
- Sirima A, Sereme A, Sereme D, Koïta K, Nana TA, et al. (2020) Effects of four essential oils on radial mycelial growth of an isolate of Alternaria sp. in Burkina Faso. Int J Biol Chem Sci 14(3): 762-771.
- Cimanga K, Kambu K, Tona L (2002) Correlation between chemical composition and antibacterial activity of essential oils of some aromatic medicinal plants growing in the Democratic Republic of Congo. J Ethnopharmacol 79(2): 213-220.
- Carlton R, Watermann R, Gray AI, Deans SG (1992) The antifungal activity of leaf gland volatile oil of sweet gale (Myrica gale) Myricaceae. Chemaecology 3: 55-59.
- Mboussi SB, Heu A, Kone Nsangou AN, Douanla Ajebe M, Ntsomboh Ntsefong G, et al. (2021) In vitro Evaluation of the Fungicidal Potential of Aqueous and Methanolic Extracts of Thevetia peruviana on the Development of Rigidoporus lignosus, Causal Agent of White Root rot of Hevea brasiliensis. Biotechnol J Int 25(1): 1-13.
-
 Table 1
Table 1 -
Table 2
-
Table 3
-
Table 4
-
Table 5
-
Table 6












QUICK LINKS
- SUBMIT MANUSCRIPT
- RECOMMEND THE JOURNAL
-
SUBSCRIBE FOR ALERTS
RELATED JOURNALS
- Journal of Infectious Diseases and Research (ISSN: 2688-6537)
- Archive of Obstetrics Gynecology and Reproductive Medicine (ISSN:2640-2297)
- Journal of Allergy Research (ISSN:2642-326X)
- Advance Research on Alzheimers and Parkinsons Disease
- Journal of Otolaryngology and Neurotology Research(ISSN:2641-6956)
- Journal of Psychiatry and Psychology Research (ISSN:2640-6136)
- International Journal of Medical and Clinical Imaging (ISSN:2573-1084)


