752
Views & Citations10
Likes & Shares
Bipolar disorder is a complex and highly heritable psychiatric disorder characterized by severe mood alterations. The precise genetic underpinnings of the disease have not been identified so far, despite numerous genome-wide association findings. This review describes the current state of genetic studies based on next generation sequencing technologies including whole exome and whole genome sequencing, as well as RNA-sequencing and highlights the fact that the integration of these studies can reveal novel knowledge such as the functional role of gene variants. However, due to the complexity of bipolar disorder, it is a compelling candidate for studies beyond DNA and RNA sequencing. Epigenetic alterations, defined as heritable but reversible modifications including DNA methylation, DNA hydroxymethylation, histone modifications and non-coding RNAs may be the link between genome and environment interactions. Additionally, a possible source of the reported immune activation in bipolar disorder is the micro biome of gastrointestinal tract, due to recent studies that indicate its pivotal role in brain function through the ‘gut-brain’ axis. The identification of methods able to modulate the micro biome emerges as a promising path for novel diagnostic and treatment options in bipolar disorder, thus the number of metagenomic studies in bipolar disorder has substantially increased the last years. Overall, the paper aims to review the most recent literature on genomic, epigenomic and metagenomic studies that have contributed to our understanding of the pathophysiology of bipolar disorder so far. The paper also focuses on the exploitation of recent advancements in high-throughput technologies for the elucidation of bipolar disorder through different approaches that may provide complementary knowledge and concludes to the need for merging the gap between all the gathered knowledge from the analysis of high-throughput data.
Keywords: Bipolar disorder, Whole exome sequencing, Whole genome sequencing, Metagenomics, Epigenomics, RNA sequencing
Abbreviations: BD: Bipolar Disorder; BDNF: Brain-Derived Neurotropic Factor; CNV: Copy Number Variation; GPCR: G protein-Coupled Receptor; GWAS: Genome-Wide Association Studies; HC: Healthy Controls; NGS: Next Generation Sequencing; NIMH: National Institute of Mental Health; RNA-seq: RNA sequencing; SZ: Schizophrenia; SNP: Single Nucleotide Polymorphism; WES: Whole Exome Sequencing; WGS: Whole Genome Sequencing
INTRODUCTION
The last twenty years have changed perspective regarding psychiatric research internationally, due to many breakthroughs in the field of genetics and high-throughput technologies that have enabled parallel sequencing of hundreds of DNA and RNA molecules. In the 20th century, family, twin and adoption studies have shown that psychiatric disorders occur in families with an established hereditary component [1]. Genome-wide association studies (GWAS) can be considered a turning point in the study of the genetic basis in Psychiatric Diseases. GWAS studies are unbiased genome screens that search for associations of common genetic variants between individuals with specific diseases. Common genetic variation in psychiatry has been proved insufficient for clinical applications, since there are still unidentified genetic factors that contribute to the diseases’ heritability, a fact that is also known as the “missing heritability” [2]. The scientific community has identified the need to integrate various lines of research into the molecular basis of such complex diseases, exploiting advancements in next generation sequencing (NGS) technologies [3].
Βipolar disorder as a heritable disorder
Bipolar disorder (BD) is a severe psychiatric disorder, characterized by extreme fluctuations in mood. The disease affects around 1% of the world’s population and it is among the main causes of disability in young people [4]. Its diagnosis is presently based on subjective clinical criteria, determined by widely acknowledged classification tools, like the Diagnostic and Statistical Manual of Mental Disorders (DSM-5) [5] or the 10th revision of the International Classification of Diseases (ICD-10) [6]. Most BD patients are classified either in BD 1 subtype (manic and/or mixed episodes) or in BD 2 subtype (depressed and/or hypomanic episodes).
Even though environmental factors can trigger BD onset, the genetic predisposition of the disease is strongly supported by family and twin studies [7].
Heritability of BD considers the percentage of genetic variation that is considered responsible for the manifestation of the disorder. Twin studies in BD estimate a heritability of around 80%. Various GWAS have identified association of numerous single nucleotide polymorphisms (SNPs), which are single-base pair alterations in a DNA sequence, at a specific DNA position in the genome of an individual [8-11]. However, even in large population studies, no consistent association of these variants with BD has been established [12-14].
Exploiting the catalytic advancements in the field of sequencing technologies, numerous studies have tried to elucidate the underlying molecular mechanisms of BD. However, much research is still required to document the main genetic and epigenetic contributors to BD.
Several approaches are encountered in the study of Bipolar Disorder. Some scientists examine the disease as a homogenous research concept, seeking for trait markers and for the elucidation of the pathophysiology of BD. Other approaches include the division and the comparison of BD 1 and BD 2 entities, while others focus on markers related to diverse emotional states of the disease (manic, mixed, depressive). Finally, there are research strategies focusing on the psychotic component of the disease through studies on BD and schizophrenic patients. These studies are usually performed on postmortem brain tissues, since the main pathophysiological site of psychiatric disorders is the brain. Other studies include cell models, such as pluripotent stem cells, immortalized peripheral leukocytes, peripheral blood leukocytes, induced neuronal cells and human skin fibroblast cells, considering that brain biopsies are not preferable in psychiatric research [15].
The paper aims to review the literature concerning the contribution of recent genetic, epigenetic and metagenomic studies.
NEXT GENERATION SEQUENCING-GENETIC STUDIES
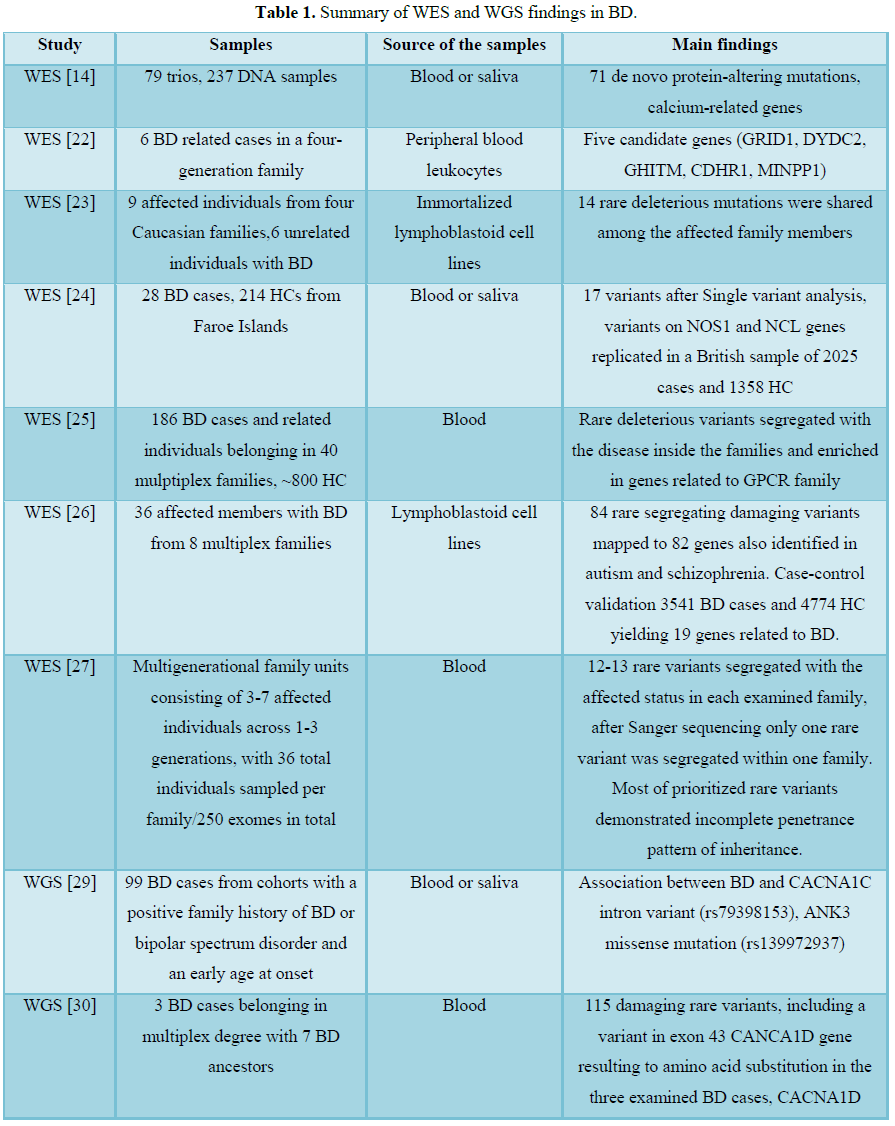
NGS technologies are high throughput technologies that perform massively parallel sequencing of base pairs in RNA and DNA samples. NGS studies in BD include whole genome sequencing (WGS) or whole exome sequencing (WES) experiments. WGS examines 98% of the whole human genome, whereas WES covers 1-2% of the human genome, including 95% of genome regions that encode a protein. The research in this field includes large Amish bipolar pedigrees provided by the National Institute of Mental Health (NIMH) Human Genetics Initiative, case-control as well as trio-based studies. Table 1 summarizes recent WES and WGS studies of BD [16].
Whole exome sequencing
Only one trio-based (case-parent study design to detect underlying variants in complex disorders) exome sequencing study has been conducted so far, including 237 samples [14]. This study revealed 71 de novo mutations also described as genetic alterations occurring in a family member for the first time due to mutations either in germ cells of the parents or in fertilized eggs. Many of the identified variants were mapped on calcium related genes. Interestingly, de novo variants with protein-altering effects occurred more frequently in BD probands with early onset of the disease. The findings support a hypothesis suggesting a relation between the higher frequencies of de novo point mutations as the paternal age advances, with the increased risk for BD in the offspring with advancing paternal age [17-21].
Another recent study investigated susceptible variants related to BD, through WES in combination with linkage analysis of 6 cases that were relatives in a four-generation family. The study resulted to five candidate genes (GRID1, DYDC2, GHITM, CDHR1, MINPP1) possibly incriminated for the onset of BD in this family [22]. Rao et al. [23] examined if rare mutations (alternative forms of a gene with a frequency in the general population usually less than 1%) located in gene-coding regions identified from WES, on immortalized lymphoblastoid cell lines of patients affected by BD, are transmitted from generation to generation and are shared between affected family members. The study resulted to 14 rare, mutations shared among the affected members of the family, with increased susceptibility for diseases (deleterious variants). The mutations were not identified in 5090 exomes of SZ patients, indicating that these mutations are specific for BD. In a study of Lescai et al. [24], exome sequencing of 28 patients affected with BD and 214 healthy controls (HC) from Faroe Islands was performed, followed by validation of the results on a sample of 2025 BD cases and 1358 HC from Britain. Sixteen mutated genes were found, among whichNOS1 and NCL were found to be replicated in the British sample [24]. In another study from Cruceanu et al. [25], exome sequencing, on blood from 186 cases and around 800 HC belonging in 40 multiplex families, was performed. The study resulted in deleterious rare variants that were inherited with the disease inside the families and interestingly were enriched in genes related to G protein-coupled receptor (GPCR) family. The genes belonging to this family of receptors are included in main drug targets [25]. Goes et al. [26] performed WES in 36 affected members with BD from 8 multiplex families, resulting to 84 rare deleterious variants. These variants were mapped to 82 genes that were identified in autism and SZ related studies, indicating an overlap of variants on risk genes for autism and SZ with rare, variants in BD families. A case-control validation followed on independent 3541 BD cases and 4774 HC yielding 19 genes related to BD [26]. A family-based exome sequencing approach was applied on blood of multigeneration families with members affected by BD. The study focused on BD cases with good response to lithium monotherapy, reducing in this way the phenotypic heterogeneity. The resulting rare variants shared among the affected members in each family, were on average twelve to thirteen. The study resulted to only one rare variant that was shared in BD patients of one affected family and was not present in the unaffected members. The findings of the study indicated a highly complex inheritance model of BD [27].
Whole genome sequencing
WGS of 99 subjects with BD resulted to a great number of variants, but the study focused on variants of genes ANK3 and CACNA1C, since they are the best-replicated genes associated with BD according to GWAS [8,11,28]. The study finally identified an association between BD and the CACNA1C variant rs79398153 located in a non-coding region of the genome, as well as the ANK3 mutation (rs139972937) [29]. In a WGS study on three cases affected by BD, 115 damaging rare variants were identified, including a variant in exon 43 of the gene CANCA1D that resulted to amino acid substitution. The three cases belonged to a multiplex pedigree with 7 BD ancestors [30]. In a WGS study performed on 200 samples from 41 families with BD, mutations were identified in genes related to neuronal excitability such as γ-aminobutyric acid and calcium channel signaling were identified [7]. Georgi et al. [31] performed linkage analysis in an Amish pedigree with members affected by BD. WGS was performed in 50 members, 23 of which were BD patients. Among the identified variants present in BD cases, rs113270504 in the gene GPR124 was of special interest since it plays a central role in angiogenesis of the central nervous system and in the blood-brain barrier development [31].
RNA SEQUENCING
Whole transcriptome studies complement our understanding regarding disease related mechanisms, through the measurement of mRNA expression levels of genes in various tissues. Over the last decade the costs of sequencing have dramatically dropped, transforming RNA Sequencing (RNA-Seq) to a commodity for dense, genetic research [32]. RNA-Seq allows the accurate quantification of gene expression levels and the analysis of the whole transcriptome for a deeper understanding of the genetic mechanisms of psychiatric disorders [33]. Transcriptomic sequencing studies act complementary to WGSand WES, deriving information about splicing events, or about the functional role of the mutated genes. However, there are factors that should be taken into consideration when examining RNA-seq studies, such as: experimental protocols performed on human brain tissues, RNA-Seq assays, and downstream bioinformatics analysis that present many differences in the various performed studies [33]. In this review an explicit account of RNA-Seq studies is aimed, since this approach has gained increasing popularity and has replaced microarrays, becoming the first option for high-throughput quantification of the whole transcriptome that overcomes limitations of microarrays technology [34].
RNA-seq studies in various cell types
A reported RNA-seq study on lymphoblastoid cell lines of BD, examined the gene expression variations that may be related to lithium response, seeking for biomarkers based on the treatment outcome. RNA-seq experiments were performed on twelve lithium responsive cases and twelve lithium non-responsive cases, followed by quantitative PCR validation on two external cohorts including 41 and 17 patients. The study resulted to fifty-six genes that presented different expression in the two studied cases, among which HDGFRP3 and ID2 presented higher expression in lithium responsive cases. These two genes are related to neurogenesis and HDGFRP3 is considered a neurotropic factor, suggesting the two genes as potential blood biomarkers for lithium response [35].
RNA-seq was also performed on induced pluripotent stem cells of BD subjects, resulting to abnormalities and hyper-excitability in young neurons of patients with BD. The observed hyper-excitability was reversed from lithium treatment in patients that responded positively in lithium treatment [36].
RNA-seq in postmortem brain
The first RNA-seq study was performed by Kohen et al. [37] on dentate gyrus granule cells of patients suffering from neuropsychiatric disorders including BD, indicating aberrant miR-182 signaling in the hippocampus of the patients. microRNAs are small noncoding RNA molecules suggested as a target for therapeutic interventions, since they are involved in regulatory mechanisms of neuronal physiology. There is also evidence from clinically depressed patients and from animal models that the dysregulations of miRNA expression are involved in molecular signaling pathways related to depressed state. Interestingly, antidepressants and selective serotonin reuptake inhibitors have been shown to reverse the observed miRNA dysregulations, suggesting them as a possible target for antidepressant treatment [38]. A more recent study in the dorsolateral prefrontal cortex resulted to altered expression of genes involved in neuroplasticity (PROM1, ABCG2, FLI1) and in circadian rhythm (OSBPL3, GANAB, SRSF5, RFX4) [39]. Cruceanu et al. [40] found differential expression of three genes belonging to the GPCR family (RXFP1, SSTR2 and CHRM2) in a study performed on the anterior cingulate of BD patients when compared with HC. Pacifico and Davis [41] identified disrupted immune response and oxidative phosphorylation after analysis of RNA-seq experiments in the dorsal striatum of BD patients.
Based on the similar clinic symptoms and the accumulating evidence for common genetic liabilities between SZ and BD, Zhao et al. [42] performed RNA-sequencing experiment on postmortem cingulate cortex of 82 SZ, BD and HC cases. In SZ cases 105 differentially expressed genes in comparison to HCs were identified, whereas 153 differentially expressed genes were identified in BD cases. Many of the identified genes presented common expression patterns in the two patient classes. Pathway analysis followed, but only for the genes that were concordant in their expression level in the two diseases and were associated with GWAS data. The analysis indicated that a shared interconnected pathway network of actin cytoskeleton and regulation of lysosomal function is implicated in the two neuropsychiatric diseases [42].
RNA-seq on medial frontal gyrus of postmortem brain of BD cases and HC examined the relative abundances of coding and non-coding RNAs. The study was the first to study circular RNAs and other long non-coding RNAs, due to their suggested involvement in brain development and neuronal integrity. Circular RNAs are long non-coding RNAs that participate in gene regulation by controlling the gene expression of microRNAs, and through their role as sequesters to RNA binding proteins. Twenty differentially expressed genes, ten long non-coding RNA transcripts and two circular transcripts (cNEBL and cEPHA3) were identified. The differentially expressed genes were validated through PCR and were enriched in biological processes involved in angiogenesis and vascular development. The study demonstrated that non-coding regions could have also a key role in the underlying RNA alterations in BD [43]. Kim et al. [44] analyzed RNA-seq data from the hippocampus of 15-well matched subjects of four categories: SZ, BD, major depression and HC. The results from the analysis of BD samples demonstrated abnormalities in synaptic proteins. Additionally, immune/inflammation co-expression modules were also built for each disease. No overlap was observed in the genes comprising the co-expression modules of the three studied diseases, indicating distinct immune-related abnormalities in each psychiatric disorder [44].
Transcriptional alterations were also observed in the post-mortem anterior cingulated cortex of BD and SZ cases compared to HC after RNA-seq experiments on 24 patients from each category. A reduced expression was observed particularly in neurospecific genes [45]. Another RNA-seq study was performed on 82 postmortem samples from three brain regions (orbitofrontal cortex, anterior cingulated cortex and dorsolateral prefrontal cortex) of SZ and BD cases as well as of HC. This study also examined the transcriptome of long non-coding RNAs and identified twenty differentially expressed long non-coding RNAs in the orbitofrontal cortex of BD cases compared to HC. After the application of weighted gene co-expression network analysis, it was indicated that the differentially expressed long non-coding RNAs in collaboration with other genes may differentiate the functions in different brain regions. The study also found that DNA methylation is involved in the observed dysregulation of long non-coding RNAs and may enhance psychotic symptoms [46].
EPIGENETIC STUDIES
The field of epigenetics concerns long-lasting alterations in gene expression through various regulatory mechanisms (e.g. DNA methylation, DNA hydroxymethylation, histone modifications), without involving DNA sequence changes. Epigenetic studies are complimentary to genetic analyses adding an extra layer in the etiology of psychiatric disorders. The normal function of epigenetic mechanisms, such as DNA methylation and histone modifications have a pivotal role in the normal function and development of the brain. Any dysregulations in these mechanisms could be proved harmful for the brain functionality or even causal for some diseases [47,48]. The epigenetic studies on BD that were performed the past few years, including DNA methylation, histone modifications and DNA hydroxymethylation are summarized in the following section.
Methylation is the most widely studied epigenetic mechanism in BD and is correlated with gene inactivation. A study on monozygotic twins discordant for BD was performed for identification of representative methylation differences, revealing four genome regions with different methylation patterns. The performed bisulfite sequencing (high-throughput sequencing for the detection of the methylome) resulted to a reduced methylation of PPIEL gene in the affected twin, a gene which is related to dopamine neurotransmitter and neuroendocrine systems [49]. Another study including two monozygotic twins reported promoter hypermethylation of the gene SLC6A4 in BD cases, which is a serotonin transporter gene [50]. This study is in accordance with another case-control study on postmortem brain samples that also concluded to promoter hypermethylation of SLC6A4 and down regulation of the gene’s expression [50]. A recent study after comparing the methylation status of BD subjects and HC found hypomethylation of potassium voltage-gated channel gene KCNQ3, which is involved hyperexcitability of the neurons [51]. In another paper, the methylation status of 5-HT3AR (5-hydroxytryptamine 3A) was associated with mood episodes, suicide in children with childhood trauma that have developed BD [52].
The identification of peripheral biomarkers remains a big challenge in clinical psychiatry, leading to constant studies that try to relate findings from studies on the brains tissues to patterns of the gene expression in the blood [53]. Peripheral methylation for example is a suggested approach, since it is accessible for many times [54,55]. Brain-derived neurotropic factor (BDNF) has been the focus of multiple methylation studies due to its important role in synaptic plasticity and stress response. In a study of Dell’Osso et al. [56], BDNF methylation levels in peripheral blood mononuclear cells samples of patients with BD 2were found to be increased but not in BD 1 when compared with HC. The methylation levels of the exon I promoter of BDNF were also found to be higher in BD 2 patients of study from D’addario et al. [57]. Soeiro de Souza et al. [58] showed that the global methylation profile of BD patients was not differentiated from the methylation profile of HC. Promoter hypomethylation of BDNF exon 1 in BD cases compared to major depression cases was also resulted from a study of Carlberg et al. [59]. These studies provided molecular evidence that BDNF may be used as a peripheral biomarker for different subtypes of BD, with possible pharmacological treatments through reverse of its implicated dysregulations.
DNA hydroxymethylation, another epigenetic mechanism has also been studied lately, in relation to neuropsychiatric disorders [60-63]. Histone deacetylases inhibit gene transcription through chromatin interactions. Based on this mechanism histone deacetylase inhibitors are suggested as possible pharmacological targets in the cognitive and behavioral fields [64]. Among the eleven histonedeacetylases that cause chromatin modifications, the histone deacetylase 4 was found overexpressed in contrast to 6 and 8 that were down regulated in the depressed state of BD cases, indicating complex expression patterns. Histone deacetylases were also found to increase the gene expression of cAMP response element binding protein, previously related to the pathophysiology of BD [63]. Sirtuin 1, 2 and 6, which are included in another family of deacetylases that interact with histones, presented alterations in the peripheral blood cells depending on the state of the patients suffering from BD and major depression [65].
In a postmortem brain study by Tang et al. [66] the levels of acetylated histone 3 in a pooled sample from BD and SZ cases were differentiated from that of HC. Another study on postmortem brains of BD subjects also showed elevated global histone H3 acetylation levels of the BD cases compared to HC of the same age [67]. H3K4 trimethylation has been found to be increased in postmortem brain of BD patients focusing on synapsin genes (SYN1, SYN2 and SYN3) and resulting in distinct synapsin profiles for BD and major depression cases [68].
The reported epigenetic modifications indicate possible implication of epigenetic mechanisms in the etiology of BD, but the exact involvement of these mechanisms in the pathophysiology of BD should be further dissected. It is interesting to examine how the independent epigenetic alterations could affect gene regulations that lead to BD development. Additionally, due to their reversibility, they have potential therapeutic applications. The study of epigenetic modifications may enable therapeutic interventions either through the identification of epigenetic mechanisms of mood stabilizers or through other substances that are developed and are tested in preclinical stage. For example, EpiEffectors are engineered molecules that target specific loci of the genome that cause chromatin alterations, and which have shown promising results in neuroscience [69].
GUT MICROBIOTA AND BD
The collective term that describes the whole population of the microorganisms in the human body is known as the micro biota and their genome as the micro biome [70,71]. The composition of the micro biota is developed in early life, can me modified due to dietary and other environmental exposures, but it is also genetically determined. Recent studies have constantly shown association between gut micro biota impairments (or else dysbiosis) and various diseases, such as obesity and autoimmune disorders [72,73].
Numerous studies support increased activity of the immune system in BD and in other neuropsychiatric disorders. The reason for this activation though remains unknown. One possible source for this activation is the microorganisms on the gastrointestinal tract [74,75]. Pathological conditions in gastrointestinal systems have been constantly reported as comorbidities of SZ and BD. The immune system is considered a mediator between the gut and the brain various pathways, thus participating in brain diseases such as BD [76,77]. Since gut micro biota is considered the main regulator of the gut-brain communication, several studies have shown that the micro biota and its genome may have a key role in neuropsychiatric disorders [77,78]. A new term has emerged the last years namely: “gut-brain axis”, indicating the bidirectional signaling between them [79,80]. All progress about gut-brain interactions has been intensified through the maturation of sequencing technologies in the last years. Metagenomic technologies have enabled the study of the whole genome of microorganisms in gut flora. Metatranscriptomics on the other hand have provided the capacity for the dynamic monitoring of their functions and viability [78]. In the following section the evidence for involvement of the gut micro biota in neuropsychiatric disorders and more specifically in BD are reported.
The gastrointestinal inflammation can be measured through biomarkers related to the microbial translocation. For example, for the diagnosis of Crohn’s disease, the blood is examined for presence of antibodies to the yeast Saccharomyces cerevisae, which is normally included in the human gut micro biome. The presence of these antibodies probably results from compromised gut mucosa-blood vasculature interface [81]. Following a similar approach, increased antibodies to the same yeast were identified in patients affected by BD and more specifically in patients with a recent onset [82]. Immune activation triggered from bacterial infections has been also identified in a study performed by Kohler et al. [83]. The prescription of antibiotic agents was used a measure of bacterial infection in 234 subjects hospitalized with acute mania, also suffering from BD as well as subjects hospitalized for schizophrenia, bipolar depression and major depression. The patients in mania group presented elevated antibiotic prescriptions compared to the other groups [84]. Two other studies have also reported relation between antibiotic exposure and mood disorders [85,86].
To elucidate the role of the micro biome in SZ and BD, clinical trials have been also performed to test the effects of compounds influencing the gut micro biome and the immune response on psychiatric symptoms. A trial of 6 month probiotic administration on individuals with acute mania resulted to similar inflammation scores with HC [87].
The study by Flowers et al compared the micro biota of individuals with BD that have either received or have not received atypical antipsychotics. The species diversity and more specifically Lachnospiraceae, Akkermansia and Sutterella were found to be reduced in females that received atypical antipsychotics compared to the females that were not treated with the respective medication [88].
In another recent study, the gut micro biota was associated with clinical parameters in a well-characterized cohort of subjects affected by BD. Additionally; the study examined the presence of differential abundancy in bacterial taxa between the affected cases and the HC. BD disease-related parameters of inflammation, immune response and metabolism were examined in relation to gut micro biota. The study identified associations between specific bacterial taxa and the studied BD parameters. The abundance of the bacterial taxa can be described by the Alpha-diversity. Variations in Alpha-diversity have been related to inflammatory bowel disease, anorexia nervosa and major depression. The study also concluded to a negative correlation of alpha diversity with BD illness duration, probably caused by inflammation processes [89].
A more recent study by Evans et al compared the micro biome of BD and HC samples obtained from stool samples to relate the micro biome to the disease burden. Differences were identified in the microbial communities and more specifically in the Gram-positive bacteria Faecalibacterium, suggesting therapeutic intervention through the increase of the specific bacterium in BD patients [90].
Despite the recent studies, it remains unclear whether the gut dysbiosis is causing the BD or vice versa. New preventive and treatment methods in psychiatric disorders may be discovered through the elucidation of these micro biome-related molecular mechanisms. Therapeutic applications based on modulators of gut micro biota are in experimental stage with few but promising results [79]. The micro biota has the advantage of being more “medically” accessible and can be easier modified compared to the human genome. Interestingly it is speculated that micro biota modifications can also affect the epigenome [91]. Studies for the development of modulators of the micro biome that could be served as treatment for psychiatric disorders are considered very promising, but several issues should be also considered, such as the way that the micro biome is affected by psychiatric medication [92].
CONCLUSION AND PERSPECTIVES
BD is considered a multiple-factor disease, triggered by environmental factors in individuals with a genetic susceptibility. Despite this established model for BD, the exact mechanisms for the causal interactions of genes and environment that lead to the onset of BD have not been identified. The great advances in the field of genetics have led to an exhaustive gene harvest, but many steps should be made to reveal the real effect of the genes and their interplay with the environment. Beyond the classical genetics, the recently emerged fields of epigenetics and metagenomics may be proved very helpful for unveiling the pathophysiological architecture of complex diseases such as BD and for moving forward to better understand and complement the scientific knowledge that has been gained from the genetic discipline. Many limitations in various methodological issues concerning the integration of the new knowledge gained from the distinct sources should be overcome to come to a point that all this knowledge will be translated to clinical applications for a better diagnosis and a permanent treatment of BD.
1. Collins AL, Sullivan PF (2013)
Genome-wide association studies in psychiatry: What have we learned? Br J
Psychiatry 202: 1-4.
2. Schulze TG (2010) Genetic research
into bipolar disorder: The need for a research framework that integrates
sophisticated molecular biology and clinically informed phenotype
characterization. Psychiatr Clin North Am 33: 67-82.
3. Cordell HJ (2009) Detecting
gene-gene interactions that underlie human diseases. Nat Rev Genet 10: 392-404.
4. Grande I, Berk M, Birmaher B,
Vieta E (2016) Bipolar disorder. Lancet 387: 1561-1572.
5. American Psychiatric Association
and American Psychiatric Association; DSM-5 Task Force (2013) Diagnostic and
statistical manual of mental disorders: DSM-5. 5th Edn, Washington
DC. Am Psychiatry Assoc xliv: 947.
6. World Health Organization (1993)
The ICD-10 classification of mental and behavioural disorders: Diagnostic
criteria for research. Geneva: World Health Organization xiii: 248.
7. Ament SA, Szelinger S, Glusman G,
Ashworth J, Hou L, et al. (2015) Rare variants in neuronal excitability genes
influence risk for bipolar disorder. Proc Natl Acad Sci U S A 112: 3576-3581.
8. Ferreira MA, O'Donovan MC, Meng
YA, Jones IR, Ruderfer DM, et al. (2008) Collaborative genome-wide association
analysis supports a role for ANK3 and CACNA1C in bipolar disorder. Nat Genet
40: 1056-1058.
9. Chen DT1, Jiang X, Akula N,
Shugart YY, Wendland JR, et al. (2013) Genome-wide association study
meta-analysis of European and Asian-ancestry samples identifies three novel
loci associated with bipolar disorder. Mol Psychiatry 18: 195-205.
10. Psychiatric G.C.B.D.W.G. (2011)
Large-scale genome-wide association analysis of bipolar disorder identifies a
new susceptibility locus near ODZ4. Nat Genet 43: 977-983.
11. Sklar P, Smoller JW, Fan J,
Ferreira MA, Perlis RH, et al. (2008) Whole-genome association study of bipolar
disorder. Mol Psychiatry 13: 558-569.
12. Goes FS (2016) Genetics of bipolar
disorder: Recent update and future directions. Psychiatr Clin North Am 39:
139-155.
13. Kato T (2015) Whole genome/exome
sequencing in mood and psychotic disorders. Psychiatr Clin Neurosci 69: 65-76.
14. Kataoka M, Matoba N, Sawada T,
Kazuno AA, Ishiwata M, et al. (2016) Exome sequencing for bipolar disorder
points to roles of de novo loss-of-function and protein-altering mutations. Mol
Psychiatry 21: 885-893.
15. Kálmán S, Garbett KA, Janka Z,
Mirnics K (2016) Human dermal fibroblasts in psychiatry research. Neuroscience
320: 105-121.
16. Craddock N, Sklar P (2013)
Genetics of bipolar disorder. Lancet 381: 1654-1662.
17. Kong A, Frigge ML, Masson G,
Besenbacher S, Sulem P, et al. (2012) Rate of de novo mutations and the
importance of father's age to disease risk. Nature 488: 471-475.
18. Chudal R, Gissler M, Sucksdorff D,
Lehti V, Suominen A, et al. (2014) Parental age and the risk of bipolar
disorders. Bipolar Disord 16: 624-632.
19. Frans E, MacCabe JH, Reichenberg A
(2008) Advancing paternal age and bipolar disorder. Arch Gen Psychiatry 65:
1034-1040.
20. Georgieva L, Rees E, Moran JL,
Chambert KD, Milanova V (2014) De novo CNVs in bipolar affective disorder and
schizophrenia. Hum Mol Genet 23: 6677-6683.
21. Malhotra D, McCarthy S, Michaelson
JJ, Vacic V, Burdick KE, et al. (2011) High frequencies of de novo CNVs in
bipolar disorder and schizophrenia. Neuron 72: 951-963.
22. Zhang T, Hou L, Chen DT, McMahon
FJ, Wang JC, et al. (2018) Exome sequencing of a large family identifies
potential candidate genes contributing risk to bipolar disorder. Gene 645:
119-123.
23. Rao AR, Yourshaw M, Christensen B,
Nelson SF, Kerner B (2017) Rare deleterious mutations are associated with
disease in bipolar disorder families. Mol Psychiatry 22: 1009-1014.
24. Lescai F, Als TD, Li Q, Nyegaard
M, Andorsdottir G, et al. (2017) Whole-exome sequencing of individuals from an
isolated population implicates rare risk variants in bipolar disorder. Transl
Psychiatry 7: e1034.
25. Cruceanu C, Schmouth JF,
Torres-Platas SG, Lopez JP, Ambalavanan A, et al. (2017) Rare susceptibility
variants for bipolar disorder suggest a role for G protein-coupled receptors.
Mol Psychiatry 223.
26. Goes FS, Pirooznia M, Parla JS,
Kramer M, Ghiban E, et al. (2016) Exome sequencing of familial bipolar
disorder. JAMA Psychiatry 73: 590-597.
27. Cruceanu C, Ambalavanan A,
Spiegelman D, Gauthier J, Lafrenière RG, et al. (2013) Family-based
exome-sequencing approach identifies rare susceptibility variants for
lithium-responsive bipolar disorder. Genome 56: 634-640.
28. Wellcome Trust Case Control (2007)
Genome-wide association study of 14,000 cases of seven common diseases and
3,000 shared controls. Nature 447: 661-678.
29. Fiorentino A, O'Brien NL, Locke
DP, McQuillin A, Jarram A, et al. (2014) Analysis of ANK3 and CACNA1C variants
identified in bipolar disorder whole genome sequence data. Bipolar Disord 16:
583-591.
30. Ross J, Gedvilaite E, Badner JA,
Erdman C, Baird L, et al. (2016) A rare variant in CACNA1D segregates with 7
bipolar I disorder cases in a large pedigree. Mol Neuropsychiatry 2: 145-150.
31. Georgi B, Craig D, Kember RL, Liu
W, Lindquist I, et al. (2014) Genomic view of bipolar disorder revealed by
whole genome sequencing in a genetic isolate. PLoS Genet 10: 1004229.
32. Casamassimi A, Federico A, Rienzo
M, Esposito S, Ciccodicola A, et al. (2017) Transcriptome profiling in human
diseases: New advances and perspectives. Int J Mol Sci 18: 1652.
33. Wu C, Bendriem RM, Garamszegi SP,
Song L, Lee CT (2017) RNA sequencing in post-mortem human brains of
neuropsychiatric disorders. Psychiatr Clin Neurosci 71: 663-672.
34. Mutz KO, Heilkenbrinker A, Lönne
M, Walter JG, Stahl F (2013) Transcriptome analysis using next-generation
sequencing. Curr Opin Biotechnol 24: 22-30.
35. Milanesi E, Voinsky, Hadar A,
Srouji A, Maj C (2017) RNA sequencing of bipolar disorder lymphoblastoid cell
lines implicates the neurotrophic factor HRP-3 in lithium's clinical efficacy.
World J Biol Psychiatry, pp: 1-13.
36. Mertens J, Wang QW, Kim Y, Yu DX,
Pham S, et al. (2015) Differential responses to lithium in hyperexcitable
neurons from patients with bipolar disorder. Nature 527: 95-99.
37. Kohen R, Dobra A, Tracy JH, Haugen
E (2014) Transcriptome profiling of human hippocampus dentate gyrus granule
cells in mental illness. Transl Psychiatry 4: e366.
38. Hansen KF, Obrietan K (2013)
microRNA as therapeutic targets for treatment of depression. Neuropsychiatr Dis
Treat 9: 1011-1021.
39. Akula N, Barb J, Jiang X, Wendland
JR, Choi KH, et al. (2014) RNA-sequencing of the brain transcriptome implicates
dysregulation of neuroplasticity, circadian rhythms and GTPase binding in
bipolar disorder. Mol Psychiatry 19: 1179-1185.
40. Cruceanu C, Tan PP, Rogic S, Lopez
JP, Torres-Platas SG, et al. (2015) Transcriptome sequencing of the anterior
cingulate in bipolar disorder: Dysregulation of G protein-coupled receptors. Am
J Psychiatry 172: 1131-1140.
41. Pacifico R, Davis RL (2017)
Transcriptome sequencing implicates dorsal striatum-specific gene network,
immune response and energy metabolism pathways in bipolar disorder. Mol
Psychiatry 22: 441-449.
42. Zhao Z, Xu J, Chen J, Kim S,
Reimers M, et al. (2015) Transcriptome sequencing and genome-wide association
analyses reveal lysosomal function and actin cytoskeleton remodeling in
schizophrenia and bipolar disorder. Mol Psychiatry 20: 563-572.
43. Luykx J (2018) Coding and
non-coding RNA dysregulation in bipolar disorder. bioRxiv.
44. Kim S, Hwang Y, Webster MJ, Lee D
(2016) Differential activation of immune/inflammatory response-related
co-expression modules in the hippocampus across the major psychiatric
disorders. Mol Psychiatry 21: 376-385.
45. Ramaker RC, Bowling KM, Lasseigne
BN, Hagenauer MH, Hardigan AA, et al. (2017) Post-mortem molecular profiling of
three psychiatric disorders. Genome Med 9: 72.
46. Hu J, Xu J, Pang L, Zhao H, Li F,
et al. (2016) Systematically characterizing dysfunctional long intergenic
non-coding RNAs in multiple brain regions of major psychosis. Oncotarget 7:
71087-71098.
47. Cholewa-Waclaw J, Bird A, von Schimmelmann
M, Schaefer A, Yu H, et al. (2016) The role of epigenetic mechanisms in the
regulation of gene expression in the nervous system. J Neurosci 36:
11427-11434.
48. Varela MA, Roberts TC, Wood MJ
(2013) Epigenetics and ncRNAs in brain function and disease: Mechanisms and
prospects for therapy. Neurotherapeutics 10: 621-631.
49. Kuratomi G, Iwamoto K, Bundo M,
Kusumi I, Kato N, et al. (2008) Aberrant DNA methylation associated with
bipolar disorder identified from discordant monozygotic twins. Mol Psychiatry
13: 429-441.
50. Sugawara H, Iwamoto K, Bundo M,
Ueda J, Miyauchi T, et al. (2011) Hypermethylation of serotonin transporter
gene in bipolar disorder detected by epigenome analysis of discordant
monozygotic twins. Transl Psychiatry 1: e24.
51. Kaminsky Z, Jones I, Verma R,
Saleh L, Trivedi H, et al. (2015) DNA methylation and expression of KCNQ3 in
bipolar disorder. Bipolar Disord 17: 150-159.
52. Perroud N, Zewdie S, Stenz L,
Adouan W, Bavamian S, et al. (2016) Methylation of serotonin receptor 3a in
Adhd, borderline personality and bipolar disorders: Link with severity of the
disorders and childhood maltreatment. Depress Anxiety 33: 45-55.
53. Rollins B, Martin MV, Morgan L,
Vawter MP (2010) Analysis of whole genome biomarker expression in blood and
brain. Am J Med Genet B Neuropsychiatr Genet 153B: 919-936.
54. Sananbenesi F, Fischer A (2009)
The epigenetic bottleneck of neurodegenerative and psychiatric diseases. Biol
Chem 390: 1145-1153.
55. Numata S, Ishii K, Tajima A, Iga
J, Kinoshita M, et al. (2015) Blood diagnostic biomarkers for major depressive
disorder using multiplex DNA methylation profiles: Discovery and validation.
Epigenetics 10: 135-141.
56. DellOsso B, D Addario C, Carlotta
Palazzo M, Benatti B, Camuri G, et al. (2014) Epigenetic modulation of BDNF
gene: Differences in DNA methylation between unipolar and bipolar patients. J
Affect Disord 166: 330-333.
57. D Addario C, DellOsso B, Palazzo
MC, Benatti B, Lietti L, et al. (2012) Selective DNA methylation of BDNF
promoter in bipolar disorder: Differences among patients with BDI and BDII.
Neuropsychopharmacology 37: 1647-1655.
58. Soeiro-de-Souza MG, Andreazza AC,
Carvalho AF, Machado-Vieira R, Young LT, et al. (2013) Number of manic episodes
is associated with elevated DNA oxidation in bipolar I disorder. Int J
Neuropsychopharmacol 16: 1505-1512.
59. Carlberg L, Scheibelreiter J,
Hassler MR, Schloegelhofer M, Schmoeger M, et al. (2014) Brain-derived
neurotrophic factor (BDNF)-epigenetic regulation in unipolar and bipolar
affective disorder. J Affect Disord 168: 399-406.
60. Kriaucionis S, Heintz N (2009) The
nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the
brain. Science 324: 929-930.
61. Kato T, Iwamoto K (2014)
Comprehensive DNA methylation and hydroxymethylation analysis in the human
brain and its implication in mental disorders. Neuropharmacology 80: 133-139.
62. Lister R, Mukamel EA (2015)
Turning over DNA methylation in the mind. Front Neurosci 9: 252.
63. Lu Q, Hutchins AE, Doyle CM,
Lundblad JR, Kwok RP (2003) Acetylation of cAMP-responsive element-binding
protein (CREB) by CREB-binding protein enhances CREB-dependent transcription. J
Biol Chem 278: 15727-34.
64. Machado-Vieira R, Frey BN,
Andreazza AC, Quevedo J (2015) Translational research in bipolar disorders.
Neural Plast 2015: 576978.
65. Duong A, Che Y, Ceylan D, Pinguelo
A, Andreazza AC, et al. (2016) Regulators of mitochondrial complex I activity:
A review of literature and evaluation in postmortem prefrontal cortex from
patients with bipolar disorder. Psychiatry Res 236: 148-157.
66. Tang B, Dean B, Thomas EA (2011)
Disease- and age-related changes in histone acetylation at gene promoters in
psychiatric disorders. Transl Psychiatry 1: e64.
67. Gavin DP, Kartan S, Chase K,
Grayson DR, Sharma RP (2008) Reduced baseline acetylated histone 3 levels and a
blunted response to HDAC inhibition in lymphocyte cultures from schizophrenia
subjects. Schizophr Res 103: 330-332.
68. Cruceanu C, Alda M, Nagy C,
Freemantle E, Rouleau GA, et al. (2013) H3K4 tri-methylation in synapsin genes
leads to different expression patterns in bipolar disorder and major
depression. Int J Neuropsychopharmacol 16: 289-299.
69. Kungulovski G, Jeltsch A (2016)
Epigenome editing: State of the art, concepts and perspectives. Trends Genet
32: 101-113.
70. Turnbaugh PJ, Ley RE, Hamady M, Fraser-Liggett
CM, Knight R, et al. (2007) The human microbiome project. Nature 449: 804-810.
71. Ursell LK, Metcalf JL, Parfrey LW,
Knight R (2012) Defining the human microbiome. Nutr Rev 70: S38-S44.
72. Khan MJ, Gerasimidis K, Edwards
CA, Shaikh MG (2016) Role of gut microbiota in the aetiology of obesity:
Proposed mechanisms and review of the literature. J Obes 2016: 7353642.
73. Li B, Selmi C, Tang R, Gershwin
ME, Ma X (2018) The microbiome and autoimmunity: A paradigm from the gut-liver
axis. Cell Mol Immunol 15: 595-609.
74. Rosenblat JD, Cha DS, Mansur RB,
McIntyre RS (2014) Inflamed moods: a review of the interactions between
inflammation and mood disorders. Prog Neuropsychopharmacol Biol Psychiatry 53:
23-34.
75. Rosenblat JD, McIntyre RS (2016)
Bipolar disorder and inflammation. Psychiatr Clin North Am 39: 125-137.
76. Erny D, Hrabe de Angelis AL, Prinz
M (2017) Communicating systems in the body: How microbiota and microglia
cooperate. Immunology 150: 7-15.
77. Fond G, Boukouaci W, Chevalier G,
Regnault A, Eberl G, et al. (2015) The"psychomicrobiotic": Targeting
microbiota in major psychiatric disorders: A systematic review. Pathol Biol
(Paris) 63: 35-42.
78. Dickerson F, Severance E, Yolken R
(2017) The microbiome, immunity and schizophrenia and bipolar disorder. Brain
Behav Immun 62: 46-52.
79. Mangiola F, Ianiro G, Franceschi
F, Fagiuoli S, Gasbarrini G, et al. (2016) Gut microbiota in autism and mood
disorders. World J Gastroenterol 22: 361-368.
80. Mayer EA (2011) Gut feelings: the
emerging biology of gut-brain communication. Nat Rev Neurosci 12: 453-466.
81. Desplat-Jego S, Johanet C, Escande
A, Goetz J, Fabien N, et al. (2007) Update on anti-Saccharomyces cerevisiae
antibodies, anti-nuclear associated anti-neutrophil antibodies and antibodies
to exocrine pancreas detected by indirect immunofluorescence as biomarkers in
chronic inflammatory bowel diseases: Results of a multicenter study. World J
Gastroenterol 13: 2312-2318.
82. Severance EG, Gressitt KL, Yang S,
Stallings CR, Origoni AE, et al. (2014) Seroreactive marker for inflammatory
bowel disease and associations with antibodies to dietary proteins in bipolar
disorder. Bipolar Disord 16: 230-240.
83. Kohler O, Benros ME, Nordentoft M,
Farkouh ME, Iyengar RL, et al. (2014) Effect of anti-inflammatory treatment on
depression, depressive symptoms and adverse effects: A systematic review and
meta-analysis of randomized clinical trials. JAMA Psychiatry 71: 1381-1391.
84. Yolken R, Adamos M, Katsafanas E,
Khushalani S, Origoni A, et al. (2016) Individuals hospitalized with acute
mania have increased exposure to antimicrobial medications. Bipolar Disord 18:
404-409.
85. Kohler O, Petersen L, Mors O,
Mortensen PB, Yolken RH, et al. (2017) Infections and exposure to
anti-infective agents and the risk of severe mental disorders: a nationwide
study. Acta Psychiatr Scand 135: 97-105.
86. Lurie I, Yang YX, Haynes K,
Mamtani R, Boursi B (2015) Antibiotic exposure and the risk for depression,
anxiety or psychosis: A nested case-control study. J Clin Psychiatry 76:
1522-1528.
87. Dickerson F, Stallings C, Origoni
A, Vaughan C, Katsafanas E, et al. (2013) A combined marker of inflammation in
individuals with mania. PLoS One 8: e73520.
88. Flowers SA, Evans SJ, Ward KM,
McInnis MG, Ellingrod VL (2017) Interaction between atypical antipsychotics and
the gut microbiome in a bipolar disease cohort. Pharmacotherapy 37: 261-267.
89. Painold A, Morkl S, Kashofer K,
Halwachs B, Dalkner N, et al. (2018) A step ahead: Exploring the gut microbiota
in inpatients with bipolar disorder during a depressive episode. Bipolar
Disord.
90. Evans SJ, Bassis CM, Hein R,
Assari S, Flowers SA, et al. (2017) The gut microbiome composition associates
with bipolar disorder and illness severity. J Psychiatr Res 87: 23-29.
91. Cenit MC, Sanz Y, Codoner-Franch P
(2017) Influence of gut microbiota on neuropsychiatric disorders. World J
Gastroenterol 23: 5486-5498.
92. Dinan TG, Stanton C, Cryan JF
(2013) Psychobiotics: A novel class of psychotropic. Biol Psychiatry 74:
720-726.
-
 Table 1
Table 1 -
Table 2

QUICK LINKS
- SUBMIT MANUSCRIPT
- RECOMMEND THE JOURNAL
-
SUBSCRIBE FOR ALERTS
RELATED JOURNALS
- Chemotherapy Research Journal (ISSN:2642-0236)
- Journal of Ageing and Restorative Medicine (ISSN:2637-7403)
- Journal of Rheumatology Research (ISSN:2641-6999)
- Journal of Otolaryngology and Neurotology Research(ISSN:2641-6956)
- Journal of Oral Health and Dentistry (ISSN: 2638-499X)
- International Journal of Radiography Imaging & Radiation Therapy (ISSN:2642-0392)
- Journal of Pathology and Toxicology Research


