896
Views & Citations10
Likes & Shares
IL-27 is a heterodimeric cytokine comprised of
IL-27p28 and EBI3. As a relatively new member of the IL-12 family, the
biological mechanisms associated with the role of IL-27 in the immune response
are ambiguous, displaying both proinflammatory and suppressive functions that
seem to be dependent on the disease model. A recent report demonstrates that
pharmacological blockade of IL-27p28 alleviates graft-versus-host disease
(GVHD) in mice. However, the specific role of the IL-27Rα/gp130 signaling
complex that forms the IL-27 receptor (IL-27R) on T cells has not been well
characterized in the context of allogeneic hematopoietic stem cell
transplantation (allo-HCT). Here, we demonstrate that IL-27Rα expression on T
cells exacerbates GVHD after allo-HCT, which was consistent across 3 different
MHC- mismatched murine models of allo-HCT. Expression of IL-27Rα on T cells was
required for acquisition of optimal Th1 effector function and subsequent
inhibition of Th2 and T regulatory subsets after allo-HCT. Furthermore,
administration of IL-27significantly increased mortality after allo-HCT;
suggesting that the suppressive functions linked to IL-27 in T cell responses
may be relatively modest in this model. Hence, IL-27Rα signaling on T cells
promotes the development of GVHD.
INTRODUCTION
Interleukin-27 (IL-27) is a heterodimeric
cytokine belonging to the IL-12 family. IL-27 is comprised of an IL-27p28a-chain and an EBI3b-chain and is the only member of the family that is not secreted as a
functional dimer [1,2]. As such, the receptor for IL-27is also heterodimeric
and is composed of a unique IL-27 receptor (IL-27R) component, or WSX-1, that
forms a complex with gp130 to transduce signaling [3]. Activated dendritic
cells (DCs) and monocytes serve as the primary source of p28 and EBI3 [1].
IL-27Rα is expressed in low levels on naïve T cells, but is upregulated on
effector and memory T cells [4]. The biological mechanisms associated with the
role of IL-27 in the immune response are ambiguous, displaying both
proinflammatory and suppressive functions that seem to be dependent on the disease
model.
Allogeneic hematopoietic stem cell
transplantation (allo-HCT) is an effective means by which to treat a wide
variety of diseases resulting from dysfunctional hematopoiesis; ranging from
certain immune deficiencies to severe blood diseases and cancers [5]. However,
the development of graft-versus-host disease (GVHD) remains the major cause of
morbidity and mortality after allo-HCT. Acute GVHD (aGVHD) generally occurs in
the first 100 days post allo-HCT and is a result of donor T cell recognition of
genetically disparate antigens presented by antigen presenting cells (APCs),
which subsequently leads to activation of both the innate and adaptive immune
responses against host epithelial tissues; namely the skin, lung, liver and
gastrointestinal tract (GI tract) [6].
A recent report demonstrates that
pharmacological blockade of IL-27p28 alleviates GVHD in mice [7]. However, the
specific role of the IL-27R/gp130 signaling complex that forms the IL-27 receptor
on T cells during GVHD
MATERIALS AND METHODS
Mice
C57BL/6 (B6; H-2b), BALB/c (H-2d),
B6-Ly5.2 (H-2b), B6D2F1 (B6 x DBA2) F1 (H-2b/d) were
purchased from NCI. IL-27R KO and BALB. B mice were purchased from Jackson Labs.
All animals were housed in specific pathogen-free conditions in the America
Association for Laboratory Animal Care-accredited Animal Resource Center at the
Medical University of South Carolina (MUSC). The Institutional Animal Care and
Use Committee of MUSC approved all work.
GVHD models
Using an X-RAD 320 irradiator, lethally
irradiated recipient BALB/c (650cGy), BALB. B (900cGy) or B6D2F1 (1200cGy)
recipients were transplanted with 5 × 106 T cell depleted bone
marrow (TCD-BM) alone or TCD-BM plus 1-3 × 106 WT or IL-27R KO T
cells and monitored for survival and body weight loss as previously described
[8-11]. T cells were purified from pooled spleen and lymph node cells by
negative selection to remove non-T cells including B cells, natural killer (NK)
cells, DCs, macrophages, granulocytes and erythroid cells. Briefly, non-T cells
were magnetically labeled with biotin-conjugated Abs against CD45R (B220),
CD49b (DX5), CD11b (Mac-1) and Ter-119, followed by anti-biotin MicroBeads
(Miltenyi Biotech, Auburn, CA). Isolation of T cells was achieved by negative
selection. Bone marrow (BM) was harvested from tibia and femurs and T cells
were depleted through complement lysis of Thy1.2+ cells.
For experiments involving adenoviral
production of IL-27, mice were injected intramuscularly with 2 × 1011
DRP of vectors 7 days prior to BMT with either vector control or IL-27AAV [12].
Flow cytometry and intracellular cytokine staining
Mononuclear cells were isolated from
recipient spleen or liver as previously described and stained for surface
markers and intracellular cytokines using standard flow cytometric protocols
[10,11]. Stained cells were analyzed using FACSDiva software, LSR II (BD
Biosciences, San Jose, CA) and FlowJo (Tree Star, Ashland, OR).The following
Abs were used for cell-surface staining: anti-CD4-V450, -APC and -PEcy7 (BD
Biosciences), anti-CD8-PEcy5, -APCcy7 and -AF700 (BD Biosciences,); anti-CD45.1-FITC
and -APC (BD Biosciences). Intracellular staining was carried out using anti-IFN-γ-PE
or Per-cp 5.5 (XMG1.2; BD Biosciences), anti-IL-4-PE (11B11; BD Pharmingen),
anti-IL-5-PE (TRFK5; BD Pharmingen), anti-Foxp3-PE (FJK-16s; eBioscience).
Statistics
For comparison of recipient survival among
groups in GVHD experiments, the log-rank test was used to determine statistical
significance. To compare body weight changes and cytokine levels, a Student t
test was performed.
RESULTS
IL-27R is required for T cells to induce GVHD
Given the recent findings that IL-27p28
exacerbates graft-versus-host disease (GVHD), we hypothesized that targeting
the alpha receptor subunit of the IL-27 receptor (IL-27Rα) specifically on T
cells would result in a reduction in GVHD severity after allogeneic bone marrow
transplantation (allo-BMT). In order to decipher the role of IL-27Rα on T
cells, we initially tested the ability of IL-27R deficient T cells to cause
GVHD in a MHC-matched but minor histocompatibility antigen (miHA) mismatched
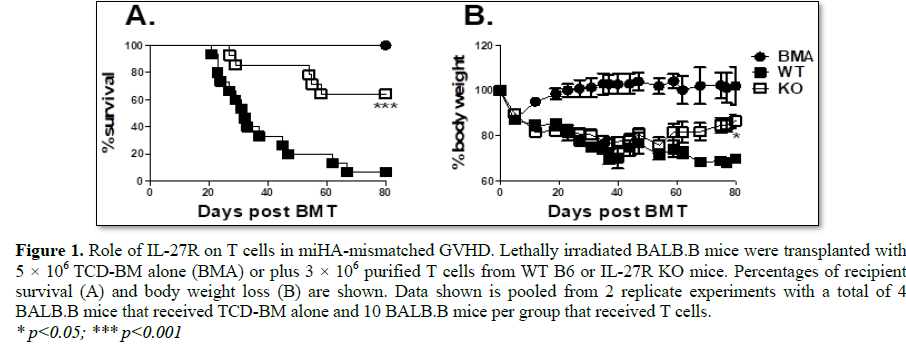
murine BMT model, C57BL/6 to BALB.B. Recipients that received T cells deficient
for IL-27Rα developed less severe GVHD, as shown by a significantly higher
survival percentage across experiments compared to WT controls (Figure 1A);
which correlated with significantly improved body weight maintenance among
groups that received IL-27Rα deficient T cells (Figure 1B). Hence, T
cells deficient for IL-27Rαhave a compromised ability to induce GVHD in a
MHC-matched model of allo-BMT.
IL-27R signaling augments Th1 responses
IL-27 was initially reported to be involved
in Th1 differentiation [1]. Therefore, we hypothesized that a decrease in IFNγ
production by T cells might be responsible for the alleviated GVHD burden seen
in the MHC-matched BMT model (Figures 1A and 1B). Consistent with this
hypothesis, we found a significant decrease in the percentage of IFNγ+ T cells
in the spleen (Figures 2A and 2C) and liver (Figures 2B and 2D)
of cohorts that received IL-27RαKO T cells 21 days’ post BMT. This data
demonstrates that IL-27Rα plays a role in T cell pathogenesis during GVHD
development and that this pathogenicity is, at least in part, attributable to
an IL-27Rα-dependent Th1 effector response.
IL-27R is required for optimal T cell pathogenicity across multiple BMT
models
Since we observed that CD4+ T helper cell
differentiation was significantly altered in IL-27R deficient T cells after
allo-BMT in the B6 to BALB.B model, we hypothesized that a similar reduction in
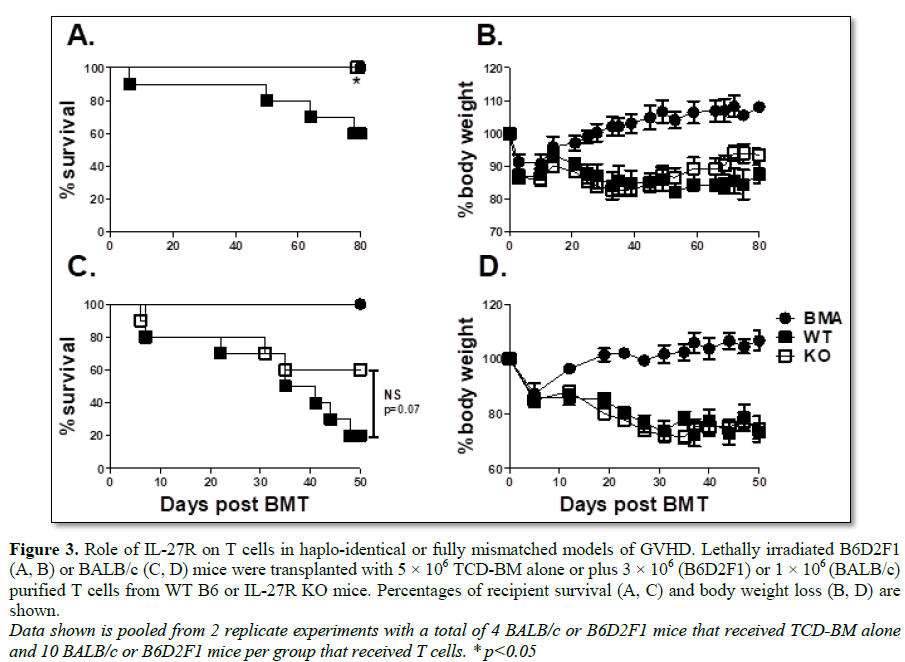
GVHD may also hold true in additional models of allo-BMT. To address this
hypothesis, we used both the B6 to B6D2F1 (Figures 3A and 3B) haplo
identical BMT model as well the B6 to BALB/c (Figures 3C and 3D) full
MHC-mismatched model. In both models, we found an increase in survival
percentage (Figures 3A and 3C) among recipients of IL-27R KO T cells
compared to WT controls after allo-BMT, albeit not statistically significant in
BALB/c recipients. This data confirms that IL-27R expression on T cells plays a
pathogenic role in GVHD development across multiple murine BMT models.
IL-27R expression on T cells inhibits differentiation toward Th2 and T
regulatory subsets after allo-BMT
Our previous results indicate that IL-27R
expression on T cells can promote GVHD after allo-BMT and that this is
attributable to an increased Th1 response. In order to corroborate this
mechanism by which IL-27R on T cells influences the development of GVHD, we
analyzed T cell proliferation and differentiation in the spleen and liver of
BALB/c recipients 21 days’ post BMT (Figure 4). In the spleen, we saw a
significant decrease in the percentage of IFNγ produced by CD4+ T cells in
recipients that received IL-27R KO T cells compared to WT controls.
Additionally, IL-27R KO CD4+ T cells isolated from the spleen produced a
significantly higher percentage of IL-4/5 and had a significantly increased
percentage of Foxp3 expression (Figures 4A and 4B). Consistently,
recipients of IL-27R KO T cells had a significantly higher percentage of
CD4+IL-4/5+ T cells in the liver (Figures 4C and 4D). These data
indicate that T cells deficient for IL-27R are skewed away from Th1
differentiation and instead differentiate intoTh2 and T regulatory subsets, a
pattern which would be manifested by reduced GVHD. However, we also noted
significant increases, albeit among few cells, in IL-17 production by IL-27R KO
CD4+ T cells in both the spleen (Figures 4A and 4B) and the liver (Figures
4C and 4D).
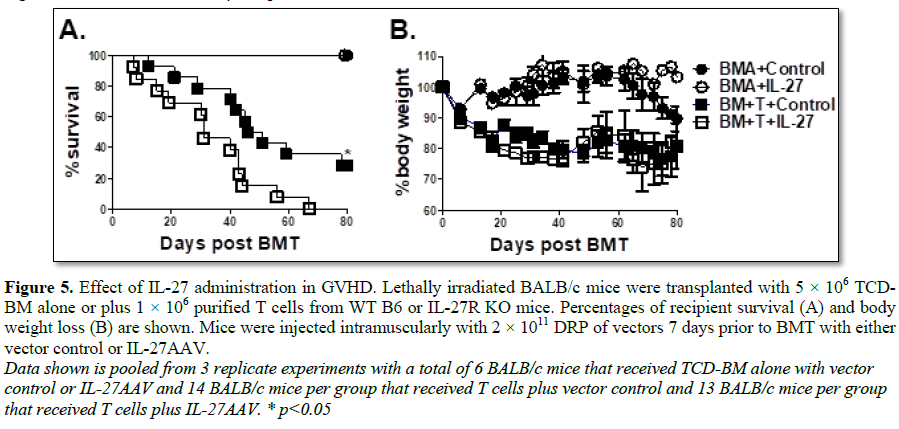
Administration of IL-27 exacerbates GVHD
Our results specifically demonstrate that
IL-27R expression on T cells augments GVHD. Taken together with previous
literature implicating IL-27p28 as a proinflammatory mediator of GVHD
development, we sought to delineate whether the cytokine IL-27 could augment T
cell mediated GVHD. To address this question, we injected either an adenoviral
vector expressing the IL-27 heterodimer (IL-27AAV) or empty vector into BALB/c
recipients 7 days prior to BMT. Lethally irradiated recipient mice were then
transplanted with B6 T cells and TCD-BM as described in Figure 1 and
monitored for survival (Figure 5A) and body weight loss (Figure 5B).
Recipient mice that were treated with IL-27AAV had significantly higher
mortality than those that received empty vector (Figure 5A); providing
further support for the notion that IL-27 signaling on T cells promotes the
development of GVHD.
DISCUSSION
In the context of autoimmunity, IL-27 has
been implicated in both pro- and anti-inflammatory responses. Taken together
with the recent report advocating for pharmacological blockade of p28 as a
potential therapy for GVHD, our results unambiguously demonstrate that IL-27
signaling on T cells exacerbates GVHD after allo-HCT.
In this study, we observed a consistent
increase in GVHD severity; not only in a MHC-matched model of allo-BMT, but
also in MHC complete-mismatched as well MHC haploidentical BMT model in cohorts
that received T cells expressing IL-27R. Mechanistically, we observed that Th1
responses were augmented in cohorts that received IL-27R competent T cells,
while Th2 and Treg differentiation were significantly decreased, consistent
with previous reports. Of note, the observed increase in IL-4/5 production by
donor T cells was quite dramatic. This is supported by previous studies which
implicate IL-27 signaling as a critical regulator of T-bet and IL-12Rβ2
expression in T cells and further demonstrate that these Th1-promoting factors
consequently suppress the master Th2 transcription factor, GATA3 [13].
Our results indicate that IL-17 production
was significantly decreased in mice that receivedIL-27R expressing T cells,
which could be a potential explanation for why we did not observe a significant
difference in body weight maintenance throughout our experiments. The increase
in IL-17 production by IL-27R deficient T cells could be potentially explained
by the significant reduction in IFNγ production, which has been reported to
negatively regulate Th17 differentiation and, hence, IL-17 production [14].
Rather, the decreased function of Th1 cells, which hypothetically would
alleviate GVHD, was offset by an increased Th17 response, resulting in
Th17-mediated pathology and subsequently no difference in weight maintenance
could be observed. This is supported by reports demonstrating IL-27a signaling
negatively regulates Th17 differentiation using other models of autoimmunity
[15,16].
In addition to demonstrating that IL-27 signaling on T cells promotes the development of GVHD, we investigated the effect of IL-27 administration after allo-BMT. In these experiments, we observed that excess IL-27 significantly increased GVHD severity. Hence, our results address the role of IL-27 in GVHD in 2 different ways, and further substantiate the claim that IL-27 signaling exacerbates GVHD. In conclusion, we provide additional evidence that IL-27 signaling is detrimental in GVHD and advocate that this pathway could be a potentially efficacious therapeutic target in clinical settings.
ACKNOWLEDGEMENT
We thank Dr. Xue-feng Bai for providing
recombinant AAV vector to express IL-27. We thank the members of DLAR and the
Flow CytometryCore Facility at the Medical University of South Carolina. The
project described was supported in part by the NIH National Center for
Advancing Translational Sciences (NCATS) through Grant Numbers TL1 TR001451 and
UL1 TR001450.This work is partially supported by R01 CA169116, CA118116,
CA169116 and R21 CA192202 (to X.-Z.Yu).
1.
Pflanz S, Timans JC, Cheung J, Rosales R, Kanzler H,
et al. (2002) IL-27, a heterodimeric cytokine composed of EBI3 and p28 protein,
induces proliferation of naive CD4+ T cells. Immunity 16: 779-790.
2.
El-Behi M, Ciric B, Dai H, Yan Y, Cullimore M, et
al. (2011) The encephalitogenicity of T(H)17 cells is dependent on IL-1- and
IL-23-induced production of the cytokine GM-CSF. Nat Immunol 12: 568-575.
3.
Aparicio-Siegmund S, Garbers C (2015) The biology of
interleukin-27 reveals unique pro- and anti-inflammatory functions in immunity.
Cytokine Growth Factor Rev 26: 579-586.
4.
Villarino AV, Larkin J, Saris CJ, Caton AJ, Lucas S,
et al. (2005) Positive and negative regulation of the IL-27 receptor during
lymphoid cell activation. J Immunol 174: 7684-7691.
5.
Ferrara JL, Levine JE, Reddy P, Holler E (2009)
Graft-versus-host disease. Lancet 373: 1550-1561.
6.
MacDonald KP, Shlomchik WD, Reddy P (2013) Biology
of graft-versus-host responses: Recent insights. Biol Blood Marrow Transplant
19: S10-S14.
7.
Belle L, Agle K, Zhou V, Yin-Yuan C, Komorowski R,
et al. (2016) Blockade of interleukin 27 signaling reduces GVHD in mice by
augmenting Treg reconstitution and stabilizing FOXP3 expression. Blood.
8.
Yu Y, Wang D, Liu C, Kaosaard K, Semple K, et al.
(2011) Prevention of GVHD while sparing GVL effect by targeting Th1 and Th17
transcription factor T-bet and RORgammat in mice. Blood 118: 5011-5020.
9.
Yu Y, Cho HI, Wang D, Kaosaard K, Anasetti C, et al.
(2013) Adoptive transfer of Tc1 or Tc17 cells elicits antitumor immunity
against established melanoma through distinct mechanisms. J Immunol 190:
1873-1881.
10.
Nguyen HD, Chatterjee S, Haarberg KM, Wu Y, Bastian
D, et al. (2016) Metabolic reprogramming of alloantigen-activated T cells after
hematopoietic cell transplantation. J Clin Invest 126: 1337-1352.
11.
Wu Y, Bastian D, Schutt S, Nguyen H, Fu J, et al.
(2015) Essential role of interleukin-12/23p40 in the development of
graft-versus-host disease in mice. Biol Blood Marrow Transplant 21: 1195-1204.
12.
Zhu X, Liu Z, Liu JQ, Zhu J, Zhang J, et al. (2016)
Systemic delivery of IL-27 by an adeno-associated viral vector inhibits T
cell-mediated colitis and induces multiple inhibitory pathways in T cells. J
Leukoc Biol 100: 403-411.
13.
Lucas S, Ghilardi N, Li J, de Sauvage FJ (2003)
IL-27 regulates IL-12 responsiveness of naive CD4+ T cells through
Stat1-dependent and -independent mechanisms. Proc Natl Acad Sci USA 100:
15047-15052.
14.
Nakae S, Iwakura Y, Suto H, Galli SJ (2007)
Phenotypic differences between Th1 and Th17 cells and negative regulation of
Th1 cell differentiation by IL-17. J Leukoc Biol 81: 1258-1268.
15.
Stumhofer JS, Laurence A, Wilson EH, Huang E, Tato
CM, et al. (2006) Interleukin 27 negatively regulates the development of
interleukin 17-producing T helper cells during chronic inflammation of the
central nervous system. Nat Immunol 7: 937-945.
16.
Diveu C, McGeachy MJ, Boniface K, Stumhofer JS,
Sathe M, et al. (2009) IL-27 blocks RORc expression to inhibit lineage
commitment of Th17 cells. J Immunol 182: 5748-5756.


QUICK LINKS
- SUBMIT MANUSCRIPT
- RECOMMEND THE JOURNAL
-
SUBSCRIBE FOR ALERTS
RELATED JOURNALS
- International Journal of AIDS (ISSN: 2644-3023)
- Journal of Clinical Trials and Research (ISSN:2637-7373)
- Stem Cell Research and Therapeutics (ISSN:2474-4646)
- Oncology Clinics and Research (ISSN: 2643-055X)
- International Journal of Surgery and Invasive Procedures (ISSN:2640-0820)
- Ophthalmology Clinics and Research (ISSN:2638-115X)
- Journal of Cardiology and Diagnostics Research (ISSN:2639-4634)



